Clear Sky Science · en
Post-catalysis structures of mitochondrial complex I with ubiquinol-10 bound in the active site
How Our Cells Turn Food into Fuel
Mitochondria are often called the power stations of our cells, and one of their key machines, known as complex I, starts the process of turning the energy in our food into the molecule ATP. This study zooms in on complex I at atomic detail to see exactly what it looks like just after it has done its job, when a small fat‑like molecule called ubiquinone-10 has just been converted into its charged, energy‑rich form. Understanding this fleeting moment helps explain how our cells so efficiently capture energy and why disruptions in this machinery can contribute to diseases of the brain, muscles and heart.
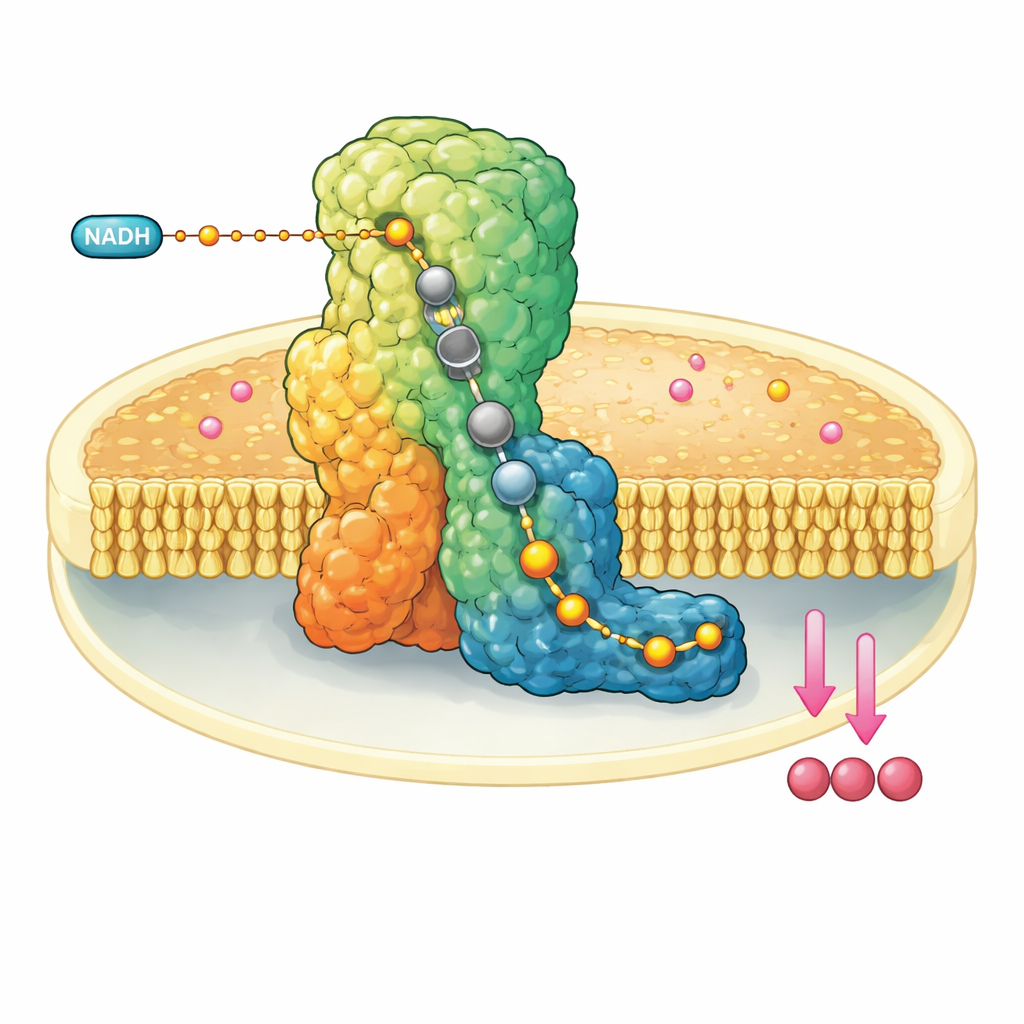
A Molecular Machine in the Mitochondrial Wall
Complex I is a huge L‑shaped protein assembly embedded in the inner membrane of mitochondria. It draws electrons from a fuel molecule, NADH, and passes them down a built‑in wire of metal clusters until they reach ubiquinone-10, a mobile carrier that shuttles electrons further along the respiratory chain. At the same time, complex I uses the released energy to move four protons across the membrane, helping build an electrochemical gradient that ultimately powers ATP synthesis. Because of its central role, malfunction of complex I is linked to a range of neuromuscular and metabolic disorders and to tissue damage during interrupted blood supply.
Freezing the Machine Mid-Breath
The researchers reconstituted bovine complex I into tiny artificial membrane discs that also contained ubiquinone-10. They then rapidly mixed in a large excess of NADH and plunge‑froze the sample within a few dozen seconds, trapping the enzyme after it had performed at least one catalytic cycle. Using state‑of‑the‑art cryo‑electron microscopy at resolutions down to about two angstroms—enough to see individual water molecules—they could reconstruct multiple three‑dimensional structures of complex I in different resting forms. Crucially, in the so‑called closed, active form, they observed ubiquinol-10, the reduced product, fully occupying the entire length of the narrow channel where the reaction takes place.
Following the Path of the Energy Carrier
In the active form, the team saw two distinct conformations of the same ubiquinol-10 molecule nestled at the end of its binding channel near the terminal iron–sulfur cluster. Compared to earlier structures with oxidized ubiquinone, the head of the molecule had shifted position and orientation, forming a strong hydrogen bond with a particular tyrosine side chain but sitting too far from a nearby histidine to interact directly. Complementary computer simulations of the surrounding charges and water molecules showed that this configuration is most stable when both that tyrosine and an aspartate residue are deprotonated—consistent with them having just donated the two protons that convert ubiquinone into ubiquinol. The second, more weakly defined pose likely represents an average over several closely related product states as ubiquinol prepares to leave the site.
Plumbing the Proton Pathways
Because the maps resolved many ordered water molecules, the authors could trace near‑continuous chains of polar residues and water through the membrane arm of complex I. These chains connect the ubiquinone reaction site to three separated “antiporter‑like” modules that move protons across the membrane. In the post‑reaction closed state with bound ubiquinol, the connections appear largely intact but punctuated by small gaps where water is likely too mobile to be seen, suggesting flexible pathways rather than rigid pipes. In contrast, an alternative open, deactive state—stabilized here by a detergent molecule lodged in the channel—shows a kinked helix that breaks one key connection, supporting the idea that this regulatory state temporarily blocks long‑range proton transfer.
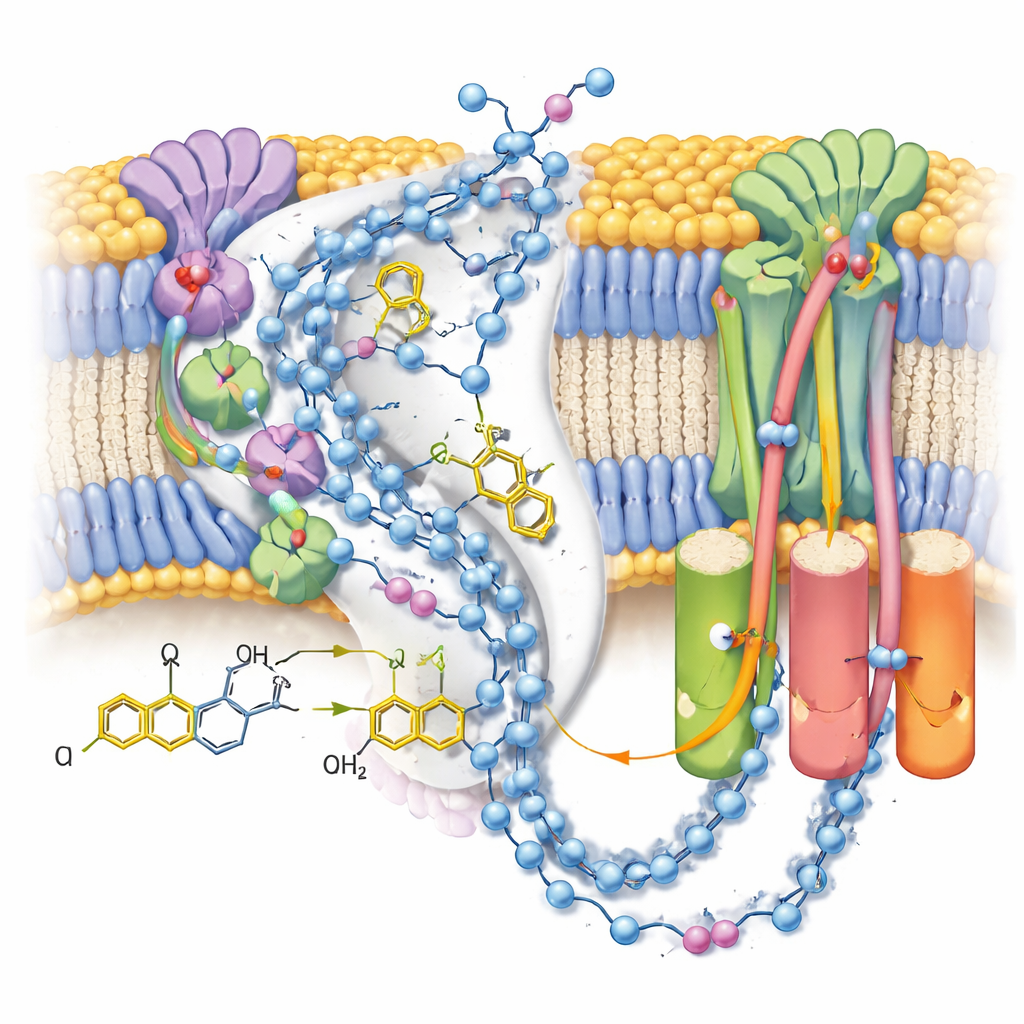
What This Means for Cell Health
Taken together, the structural snapshots and simulations support a sequence in which ubiquinone binds in a pre‑reaction pose, accepts two electrons and two protons to become ubiquinol, and then lingers in a post‑reaction pose while the nearby protein groups wait to be re‑protonated from the matrix side of the membrane. The work clarifies how subtle changes in the position and charge of a few atoms at the active site can ripple through long proton pathways to drive pumping, and how alternative, deactive forms can interrupt this flow. By sharpening our view of complex I at work, these results provide a framework for understanding inherited defects, drug action and the mitochondrial damage that occurs during events such as heart attacks and strokes.
Citation: Chung, I., Pereira, C.S., Wright, J.J. et al. Post-catalysis structures of mitochondrial complex I with ubiquinol-10 bound in the active site. Nat Commun 17, 3506 (2026). https://doi.org/10.1038/s41467-026-70030-0
Keywords: mitochondrial complex I, ubiquinone-10, cryo-EM structure, proton pumping, oxidative phosphorylation