Clear Sky Science · zh
通过释放关键结构约束使视觉G蛋白偶联受体视紫红质的暗态构象被G蛋白激活
为什么一种夜视蛋白很重要
在近黑暗环境中看见事物依赖于我们眼睛中的一种感光蛋白——视紫红质。通常,视紫红质在受到光子照射前几乎完全保持沉默,从而帮助我们避免在昏暗场景中出现会模糊视野的“视觉噪声”。但在某些遗传性眼病中,视紫红质即使在黑暗中也会过度活跃,扰乱视觉系统并损害夜间视力。本研究解析了视紫红质结构中的微小变化如何将其从紧密锁定的静止态翻转为活跃的信号态——在完全无光的情况下——从而为理解我们的视觉系统如何控制其非凡灵敏度提供了新见解。 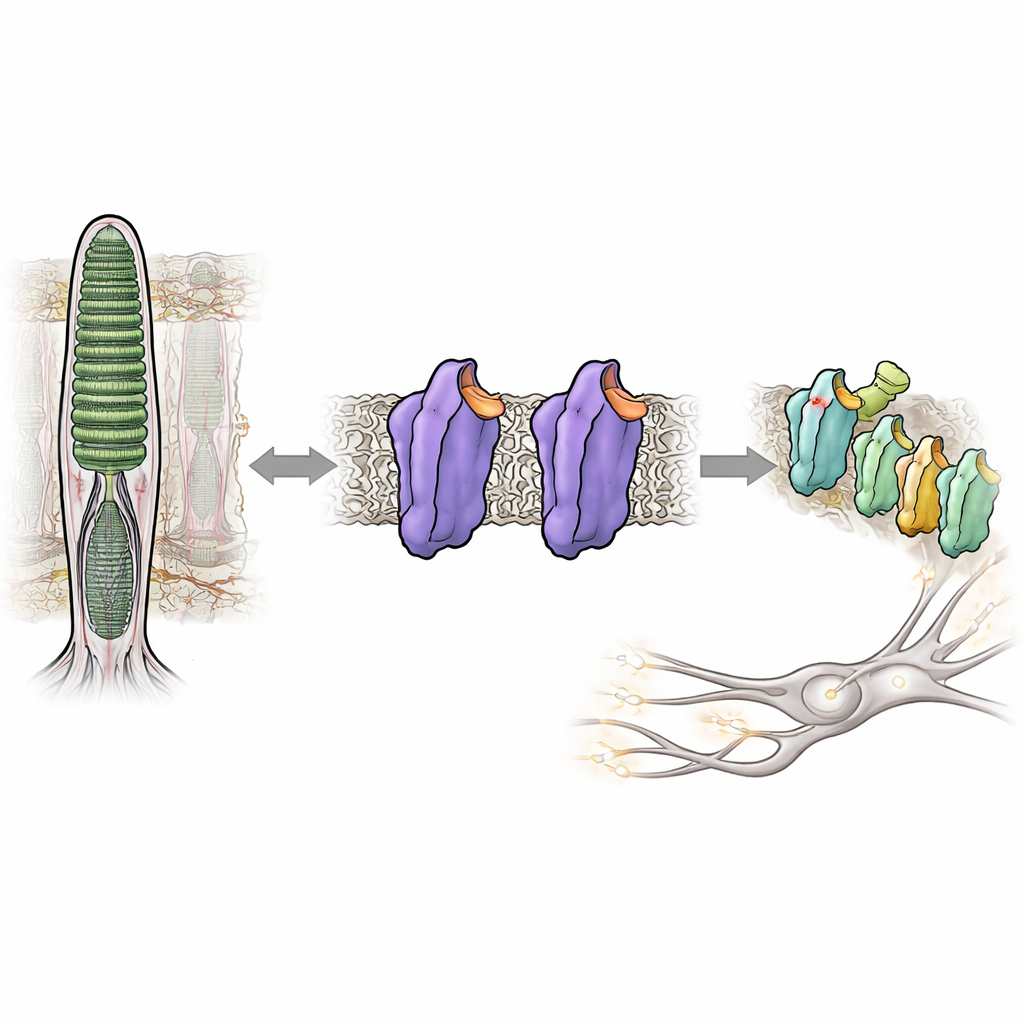
眼中的分子光开关
视紫红质属于一大类细胞开关——G 蛋白偶联受体,这类受体响应激素、气味和许多药物。在视网膜的杆细胞中,视紫红质被调谐以响应极其微弱的光线。它通过结合一种来源于维生素A的小分子11-顺视黄醛来实现,这种分子充当内在的刹车,将蛋白保持在静默或暗态。当光照射时,视黄醛瞬间改变构型,触发视紫红质的一系列结构变化,使其转变为称为 Meta II 的活性构象。该活性构象随后激活一个伴侣蛋白——称为变幅蛋白的G蛋白,启动被大脑解读为视觉的电信号。
当暗态松动时
有些人遗传到视紫红质中的微小改变或突变,这些突变削弱了这种暗态刹车,使蛋白在无光条件下也发生过多信号传导。此类“泄漏”活性与先天性固定性夜盲和色素性视网膜炎等疾病有关。作者聚焦于视紫红质内的三个位置,这些位置作为内部“微开关”发挥作用,各自有助于将蛋白保持在不活动的构象。已知这些位点的单个突变可以轻微地破坏暗态稳定性或增加自发活性。在此研究中,研究人员将它们组合成工程化的双突变和三突变,以观察同时放松几个约束是否足以在完全黑暗中将视紫红质推向类活性状态。
构建永久准备就绪的视紫红质
研究团队在培养细胞中表达突变视紫红质,对其进行纯化,并与其正常的视黄醛辅因子重组。通过紫外-可见光谱学,他们追踪了这些色素如何吸收光、在较高温度下的稳定性,以及对光照和酸性条件的响应。携带全部三处替换的三重突变体在无照射条件下显示出与活性 Meta II 形式匹配的优势吸收带。它在热力学上不稳定,迅速失去暗态的光谱特征,这与易于向活性构象转换的蛋白相一致。值得注意的是,该突变体甚至可以在黑暗中结合全反视黄醛——这是正常视紫红质无法做到的,进一步表明其构象已呈“开放”状态。功能测试证实,该三重突变体在黑暗中完全激活了G蛋白,并且在光照后其表现与正常视紫红质相当或更佳。 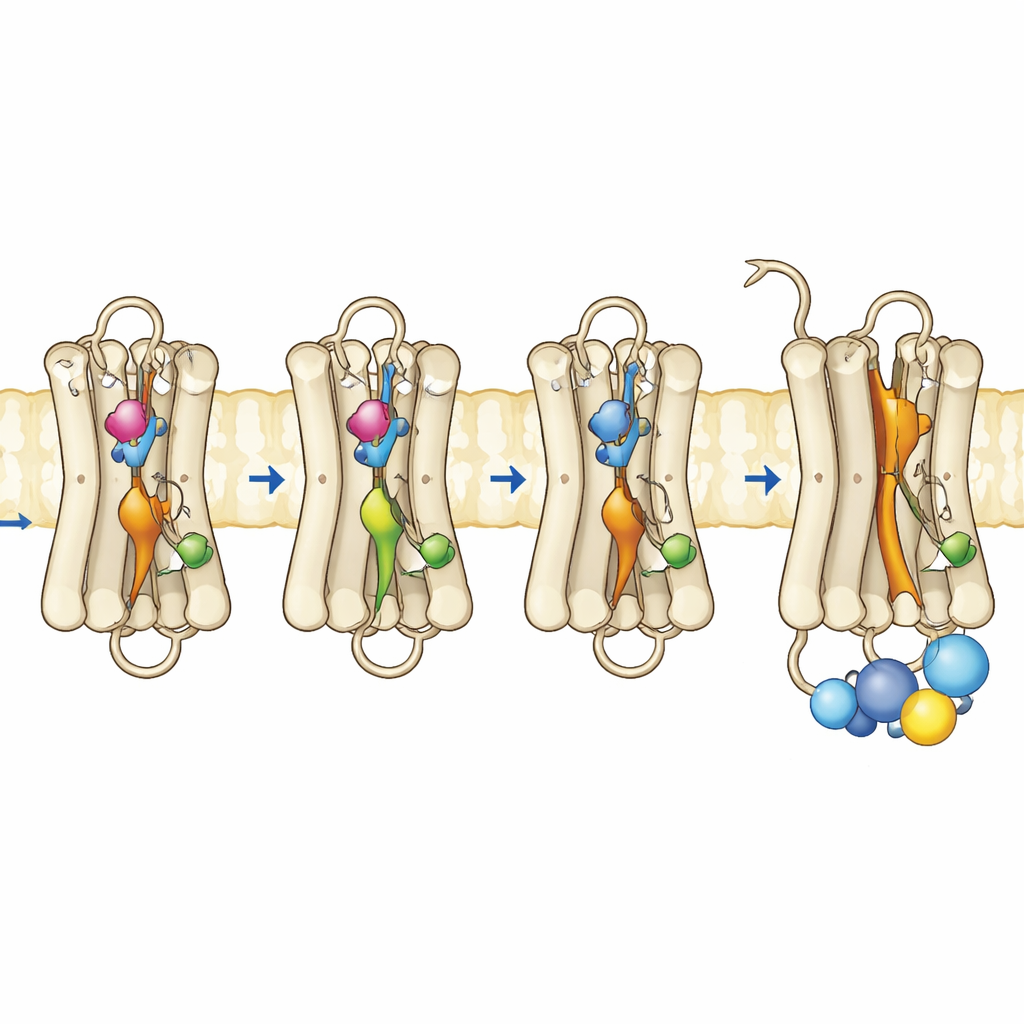
观察蛋白的移动
为了解在运动性和柔性层面发生了什么变化,作者使用基于荧光的方法来报告靠近视紫红质内表面的特定螺旋(称为第8螺旋)如何移动,以及当G蛋白的一个荧光肽片段接近受体时其行为。在正常视紫红质中,只有在光激活受体后才出现强烈的荧光变化。相比之下,三重突变体和其中一个单点突变体在黑暗中就已显示出类活性行为,表明G蛋白停靠的内表面已发生重组。时间分辨荧光各向异性测量显示,三重突变体中的第8螺旋更具可动性并占据不同的构象空间,类似于活性视紫红质。对膜中蛋白的互补计算机模拟支持了这些发现:这三处突变协同破坏了关键的内部接触,使若干内部开关在没有光引发的视黄醛构型翻转情况下也能采取类活性几何构象。
这对视力和疾病意味着什么
光谱学测量、功能测验、荧光研究和模拟共同表明,仅改变三个精心选择的位点就足以解锁视紫红质的暗态并使其自身驱动进入类活性构象。本质上,这些突变放松了通常在光到来之前保持受体静默的关键内部支撑。这项工作阐明了在200多个氨基酸中仅有少数残基如何控制这一关键视觉蛋白在沉默与信号传导之间的转换。理解这些结构尺度的杠杆有助于解释某些遗传突变如何引起过度活性和夜间视力问题,并为G蛋白偶联受体在全身范围内如何在稳定性与响应性之间取得平衡提供了更通用的蓝图。
引用: Ramon, E., Kirchberg, K., Jiménez-Rosés, M. et al. G-protein activation of the dark-state conformation of the visual G protein-coupled receptor rhodopsin by releasing critical structural constraints. Commun Biol 9, 523 (2026). https://doi.org/10.1038/s42003-026-09774-w
关键词: 视紫红质, 夜间视力, G 蛋白偶联受体, 视网膜疾病, 蛋白突变