Clear Sky Science · pl
Aktywacja białka G w ciemnym stanie konformacyjnym receptoru ródopsyny sprzężonego z białkiem G poprzez zwolnienie krytycznych ograniczeń strukturalnych
Dlaczego białko odpowiedzialne za widzenie nocne ma znaczenie
Widzenie w bliskiej ciemności zależy od czujnego na światło białka w naszych oczach, zwanego ródopsyną. Zwykle ródopsyna pozostaje niemal całkowicie uśpiona, dopóki nie trafi w nią foton, co zapobiega wizualnemu „szumowi”, który rozmywałby słabe sceny. Jednak w niektórych dziedzicznych chorobach oczu ródopsyna staje się nadmiernie aktywna nawet w ciemności, dezorganizując system wzrokowy i pogarszając widzenie nocne. W tym badaniu rozkładano na czynniki pierwsze, jak drobne zmiany struktury ródopsyny mogą przełączyć ją ze ściśle zamkniętego, cichego stanu w aktywny stan sygnalizacyjny — bez żadnego światła — dostarczając nowych informacji o tym, jak nasz system wzrokowy utrzymuje swoją niezwykłą czułość pod kontrolą. 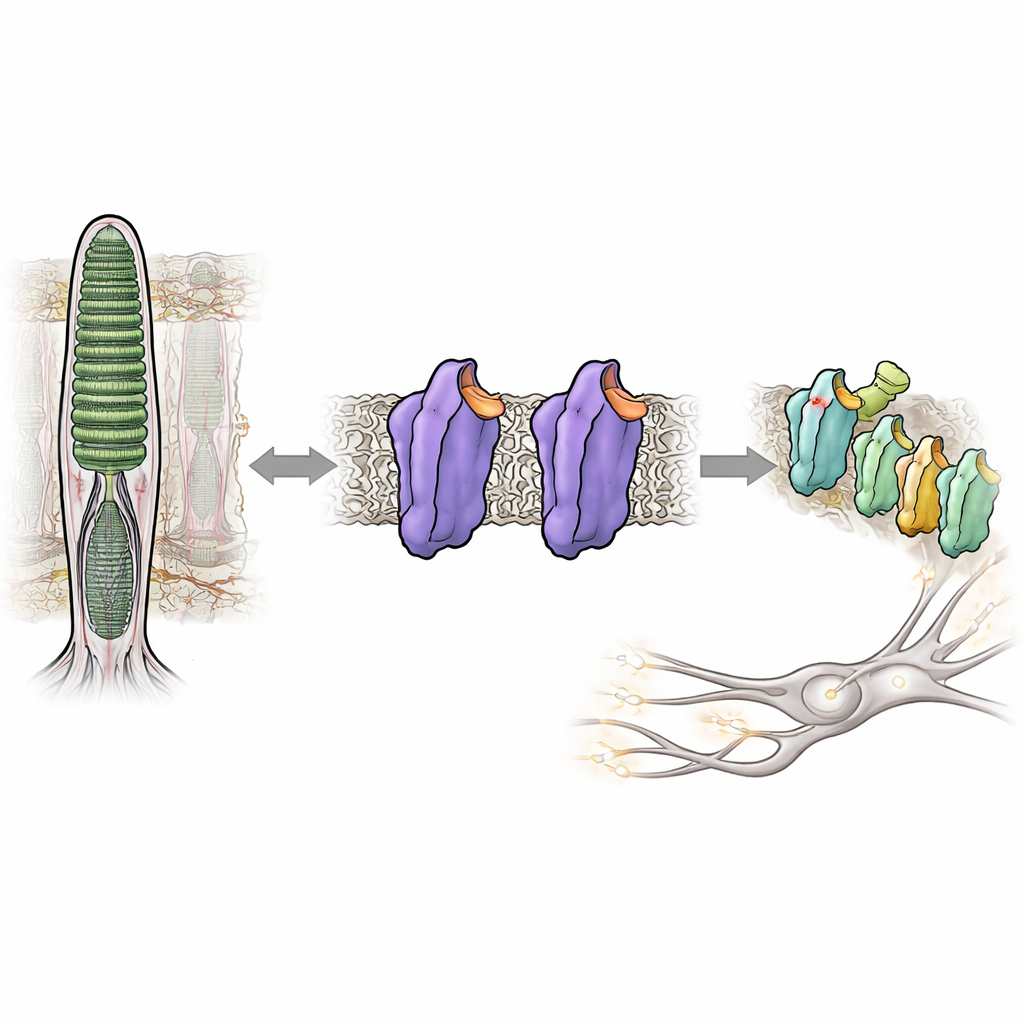
Biomolekularny wyłącznik światła w oku
Ródopsyna należy do ogromnej rodziny komórkowych przełączników zwanych receptorami sprzężonymi z białkiem G, które odpowiadają na hormony, zapachy i wiele leków. W pręcikach siatkówki ródopsyna jest dostrojona do reagowania na najdrobniejsze błyski światła. Robi to, wiążąc małą pochodną witaminy A, 11-cis-retinal, która działa jak wbudowany hamulec, utrzymując białko w cichym, czyli ciemnym, stanie. Gdy pada światło, retinal natychmiast zmienia kształt, wywołując kaskadę zmian strukturalnych w ródopsynie i przekształcając ją w aktywną formę zwaną Meta II. Ta aktywna forma następnie włącza białkowego partnera, białko G zwane transducyną, zapoczątkowując sygnał elektryczny, który mózg interpretuje jako obraz.
Gdy ciemny stan się poluzowuje
Niektórzy ludzie dziedziczą drobne zmiany, czyli mutacje, w ródopsynie, które osłabiają ten ciemny hamulec i pozwalają białku nadmiernie sygnalizować nawet bez światła. Taka „przeciekająca” aktywność wiąże się z zaburzeniami takimi jak wrodzona ustalona ślepota nocna i retinitis pigmentosa. Autorzy skupili się na trzech pozycjach w obrębie ródopsyny, które działają jak wewnętrzne „mikroprzełączniki”, każdy pomagający utrzymać białko w jego nieaktywnej formie. Pojedynczo znane mutacje w tych miejscach mogą łagodnie destabilizować stan ciemny lub zwiększać spontaniczną aktywność. W tym badaniu badacze połączyli je w inżynieryjne podwójne i potrójne mutanty, aby sprawdzić, czy jednoczesne poluzowanie kilku ograniczeń wypchnie ródopsynę w kierunku stanu przypominającego aktywność całkowicie w ciemności.
Budowanie trwale przygotowanej ródopsyny
Zespół wytworzył mutanty ródopsyny w hodowanych komórkach, oczyścił je i ponownie złożył z normalnym koenzymem retinalu. Przy użyciu spektroskopii w zakresie ultrafioletu–widzialnym śledzili, jak te barwniki absorbują światło, jak stabilne są w wyższych temperaturach oraz jak reagują na ekspozycję na światło i warunki kwaśne. Potrójny mutant, niosący wszystkie trzy substytucje, wykazywał dominujące pasmo absorpcji odpowiadające aktywnej formie Meta II, nawet bez oświetlenia. Był termicznie niestabilny, szybko tracąc spektralny znak stanu ciemnego, co zgadza się z białkiem, które chętnie przechodzi w aktywną konformację. Co znamienne, ten mutant mógł nawet wiązać all-trans-retinal — formę chromoforu aktywowaną światłem — w ciemności, czego normalna ródopsyna nie potrafi, co dodatkowo wskazuje na już „otwartą” konformację. Testy funkcjonalne potwierdziły, że ten potrójny mutant w pełni aktywował białko G w ciemności i działał tak dobrze lub lepiej niż normalna ródopsyna po ekspozycji na światło. 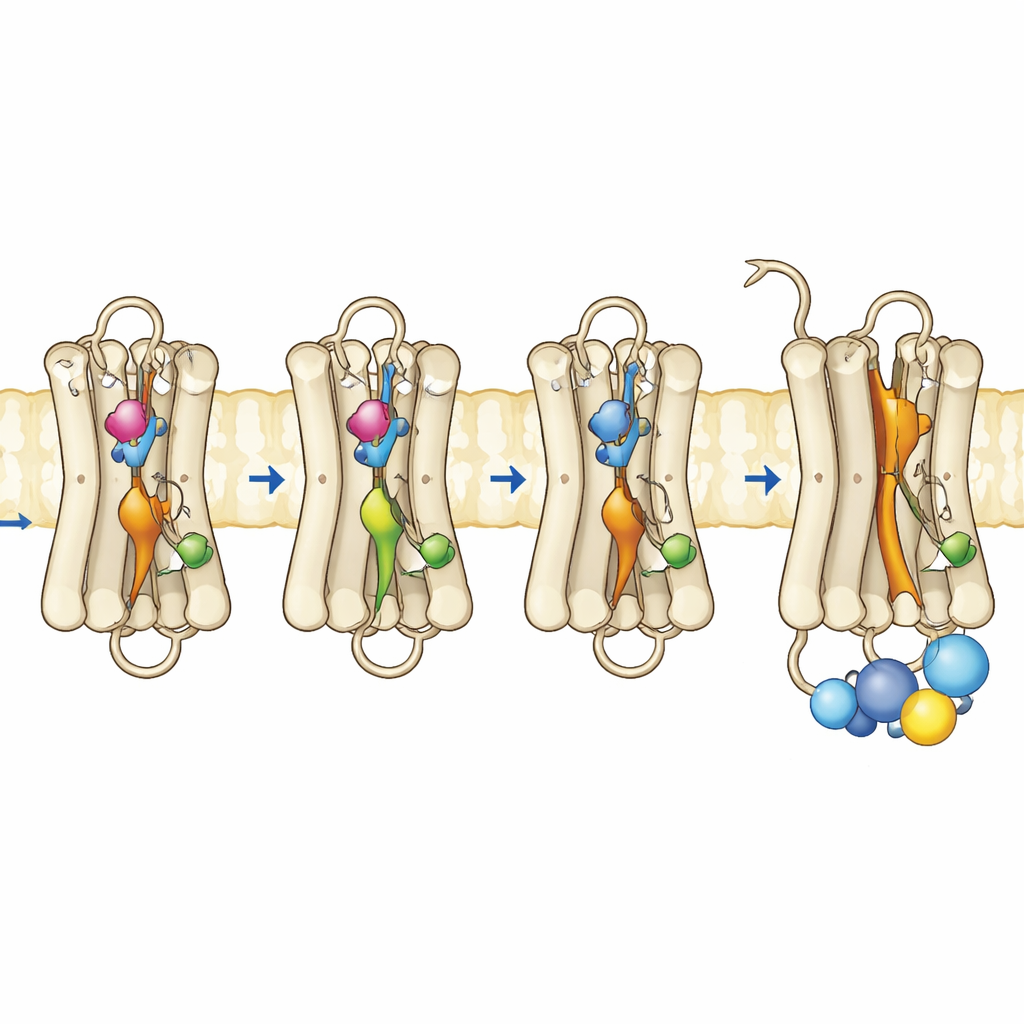
Obserwowanie ruchu białka
Aby zobaczyć, co zmieniło się na poziomie ruchu i elastyczności, autorzy zastosowali metody oparte na fluorescencji, które informują o ruchach konkretnej helisy przy wewnętrznej powierzchni ródopsyny, zwanej helisą 8, oraz o zachowaniu fluorescencyjnego fragmentu peptydowego białka G, gdy zbliża się do receptora. W normalnej ródopsynie silne zmiany fluorescencji pojawiają się dopiero po aktywacji receptora światłem. W przeciwieństwie do tego potrójny mutant i jeden z mutantów pojedynczych już w ciemności wykazywały zachowanie podobne do aktywnego, co wskazuje, że wewnętrzna powierzchnia, gdzie dokuje białko G, uległa reorganizacji. Pomiary czasowo-rozdzielczej anizotropii fluorescencji ujawniły, że helisa 8 w potrójnym mutancie była bardziej ruchoma i zajmowała inną przestrzeń konformacyjną, przypominającą tę w aktywnej ródopsynie. Uzupełniające symulacje komputerowe białka w membranie potwierdziły te ustalenia: trzy mutacje współdziałając naruszały kluczowe wewnętrzne kontakty, pozwalając kilku wewnętrznym przełącznikom przyjąć geometrię podobną do aktywnej nawet bez świetlnego przełomu retinalu.
Co to oznacza dla widzenia i chorób
Razem pomiary spektroskopowe, testy funkcjonalne, badania fluorescencyjne i symulacje pokazują, że zmiana zaledwie trzech starannie wybranych miejsc wystarcza, by odblokować ciemny stan ródopsyny i sama doprowadzić ją do konformacji przypominającej aktywną. W istocie te mutacje luzują krytyczne wewnętrzne zaczepy, które normalnie utrzymują receptor w spokoju, dopóki nie nadejdzie światło. Praca ta wyjaśnia, jak zaledwie kilka reszt z ponad 200 aminokwasów może kontrolować przejście między ciszą a sygnalizacją w kluczowym białku wzrokowym. Zrozumienie tych dźwigni strukturalnych pomaga wyjaśnić, jak niektóre dziedziczne mutacje powodują nadmierną aktywność i problemy z widzeniem nocnym, a także oferuje bardziej ogólny schemat tego, jak receptory sprzężone z białkiem G równoważą stabilność z reaktywnością w całym organizmie.
Cytowanie: Ramon, E., Kirchberg, K., Jiménez-Rosés, M. et al. G-protein activation of the dark-state conformation of the visual G protein-coupled receptor rhodopsin by releasing critical structural constraints. Commun Biol 9, 523 (2026). https://doi.org/10.1038/s42003-026-09774-w
Słowa kluczowe: ródopsyna, widzenie nocne, receptory sprzężone z białkiem G, choroby siatkówki, mutacje białek