Clear Sky Science · it
Attivazione da parte della proteina G della conformazione nello stato oscuro del recettore visivo accoppiato a proteina G rodopsina rilasciando vincoli strutturali critici
Perché una proteina per la visione notturna è importante
La capacità di vedere quasi al buio dipende da una proteina sensibile alla luce nei nostri occhi chiamata rodopsina. Normalmente la rodopsina rimane quasi completamente silente finché un fotone non la colpisce, aiutandoci a evitare il “rumore” visivo che offuscherebbe le scene deboli. Ma in alcune malattie ereditarie dell’occhio, la rodopsina diventa troppo attiva anche al buio, confondendo il sistema visivo e compromettendo la visione notturna. Questo studio analizza come piccole modifiche nella struttura della rodopsina possano trasformarla da uno stato strettamente bloccato e silenzioso in uno stato attivo e segnalante—senza alcuna luce—offrendo nuove intuizioni su come il nostro sistema visivo mantenga sotto controllo la sua straordinaria sensibilità. 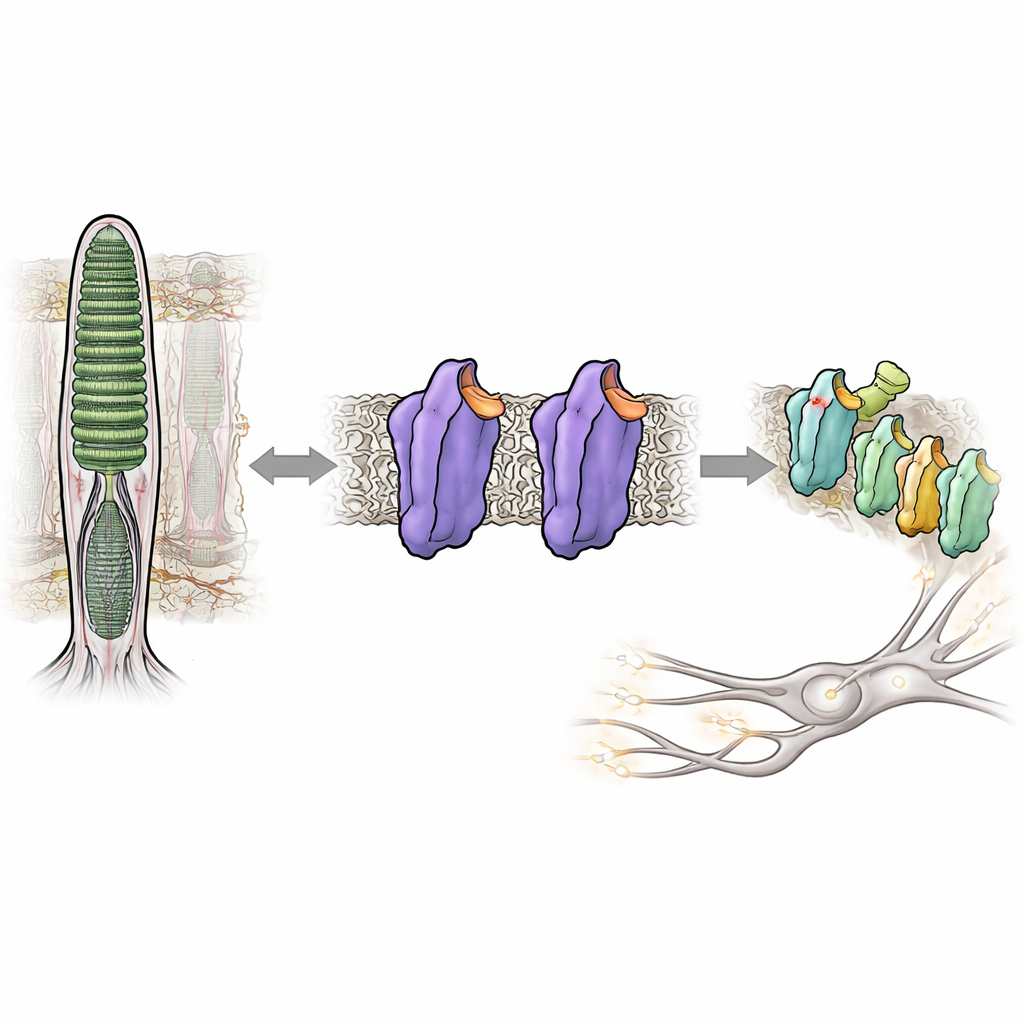
Un interruttore molecolare nell’occhio
La rodopsina appartiene a una vasta famiglia di interruttori cellulari chiamati recettori accoppiati a proteine G, che rispondono a ormoni, odori e molti farmaci. Nelle cellule bastoncellari della retina, la rodopsina è sintonizzata per rispondere al più tenue bagliore di luce. Lo fa legando una piccola molecola derivata dalla vitamina A, l’11-cis-retinale, che funge da freno incorporato, mantenendo la proteina in uno stato silenzioso o scuro. Quando la luce colpisce, il retinal subisce un cambiamento di forma che innesca una cascata di cambiamenti strutturali nella rodopsina e la trasforma in una forma attiva nota come Meta II. Questa forma attiva attiva poi una proteina partner, una proteina G chiamata transducina, avviando il segnale elettrico che il cervello interpreta come visione.
Quando lo stato oscuro si allenta
Alcune persone ereditano piccoli cambiamenti, o mutazioni, nella rodopsina che indeboliscono questo freno oscuro e permettono alla proteina di segnalare troppo anche senza luce. Tale attività “perdente” è collegata a disturbi come la cecità notturna congenita stazionaria e la retinite pigmentosa. Gli autori si sono concentrati su tre posizioni nella rodopsina che funzionano come “microswitch” interni, ciascuno contribuendo a mantenere la proteina nella sua forma inattiva. Singolarmente, mutazioni note in questi siti possono destabilizzare lievemente lo stato oscuro o aumentare l’attività spontanea. Qui i ricercatori le hanno combinate in mutanti doppi e tripli ingegnerizzati per verificare se il rilassamento simultaneo di più vincoli spingesse la rodopsina verso uno stato simile all’attivo completamente al buio.
Costruire una rodopsina permanentemente pronta
Il team ha prodotto rodopsine mutate in cellule coltivate, le ha purificate e le ha riassemblate con il loro normale cofattore retinal. Utilizzando spettroscopia nell’ultravioletto–visibile, hanno monitorato come questi pigmenti assorbivano la luce, quanto fossero stabili a temperature più elevate e come rispondevano all’esposizione alla luce e a condizioni acide. Il mutante triplo, portatore di tutte e tre le sostituzioni, ha mostrato una banda di assorbimento dominante corrispondente a quella della forma attiva Meta II, anche senza illuminazione. Era termicamente instabile, perdendo rapidamente la sua firma spettrale dello stato oscuro, coerente con una proteina che passa facilmente verso una conformazione attiva. Notevolmente, questo mutante poteva persino legare l’all-trans-retinale—la forma del cromoforo attivata dalla luce—al buio, cosa che la rodopsina normale non può fare, indicando ulteriormente una conformazione già “aperta”. I test funzionali hanno confermato che questo mutante triplo attivava completamente la proteina G al buio e funzionava tanto bene quanto, o meglio di, una rodopsina normale dopo l’esposizione alla luce. 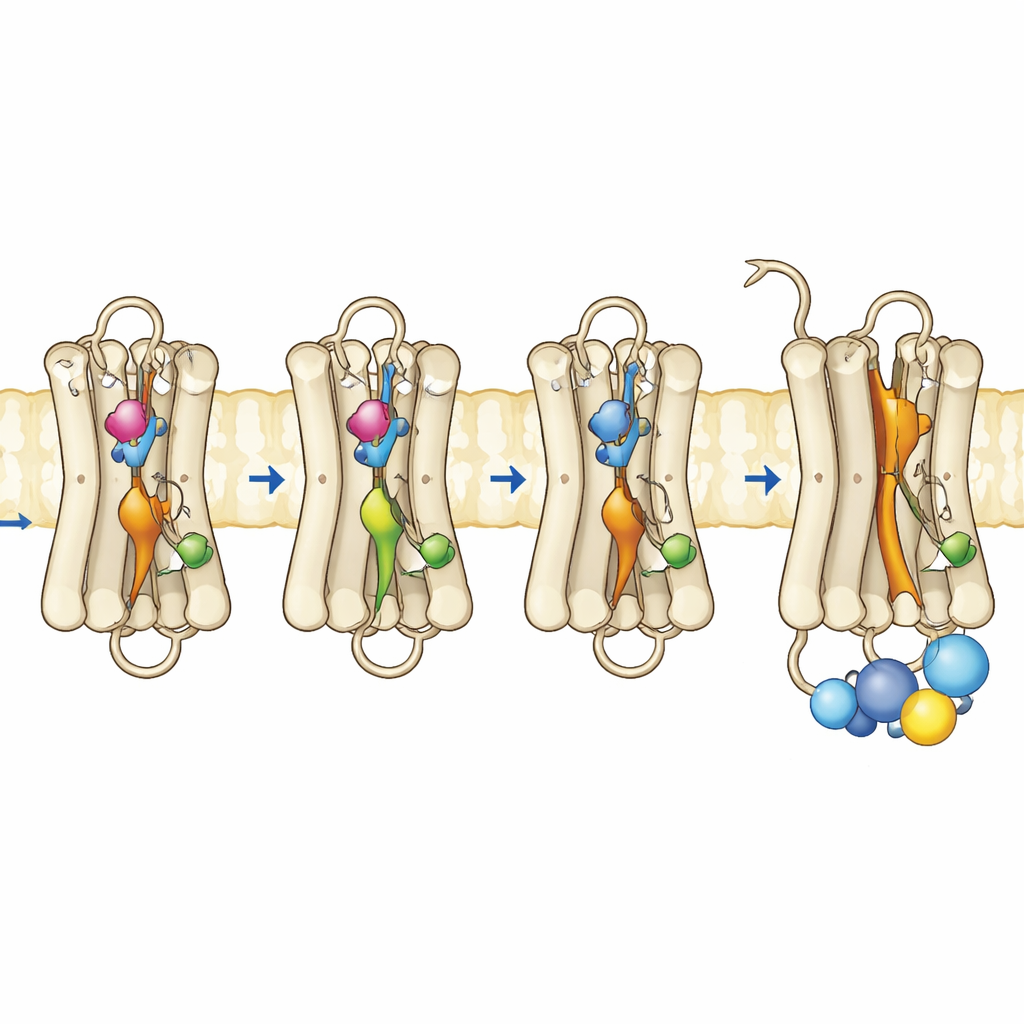
Osservare il movimento della proteina
Per vedere cosa cambiava a livello di movimento e flessibilità, gli autori hanno usato metodi basati sulla fluorescenza che riferiscono su come un’elica specifica vicino alla superficie interna della rodopsina, chiamata elica 8, si muove e su come un frammento peptidico fluorescente della proteina G si comporta quando si avvicina al recettore. Nella rodopsina normale, forti variazioni di fluorescenza compaiono solo dopo che la luce ha attivato il recettore. Al contrario, il mutante triplo e uno dei mutanti a sito singolo mostravano già un comportamento simile all’attivo nel buio, indicando che la superficie interna dove si ancora la proteina G si era riorganizzata. Misurazioni di anisotropia di fluorescenza risolta nel tempo hanno rivelato che l’elica 8 nel mutante triplo era più mobile e occupava uno spazio conformazionale diverso, somigliante a quello della rodopsina attiva. Simulazioni al computer complementari della proteina in una membrana hanno supportato questi risultati: le tre mutazioni interrompevano cooperativamente contatti interni chiave, permettendo a diversi interruttori interni di adottare geometrie simili a quelle attive anche senza il ribaltamento del retinal indotto dalla luce.
Cosa significa per la visione e la malattia
Nel complesso, le misurazioni spettroscopiche, i saggi funzionali, gli studi di fluorescenza e le simulazioni mostrano che alterare solo tre siti scelti con cura è sufficiente per sbloccare lo stato oscuro della rodopsina e guidarla autonomamente verso una conformazione simile a quella attiva. In sostanza, queste mutazioni rilassano i supporti interni critici che normalmente mantengono il recettore silente fino all’arrivo della luce. Questo lavoro chiarisce come poche residue tra le oltre 200 amminoacidi possano controllare la transizione tra silenzio e segnalazione in una proteina visiva chiave. Comprendere queste leve strutturali aiuta a spiegare come certe mutazioni ereditarie causino attività eccessiva e problemi della visione notturna, e offre uno schema più generale su come i recettori accoppiati a proteine G bilancino stabilità e reattività in tutto il corpo.
Citazione: Ramon, E., Kirchberg, K., Jiménez-Rosés, M. et al. G-protein activation of the dark-state conformation of the visual G protein-coupled receptor rhodopsin by releasing critical structural constraints. Commun Biol 9, 523 (2026). https://doi.org/10.1038/s42003-026-09774-w
Parole chiave: rodopsina, visione notturna, recettori accoppiati a proteine G, malattie retiniche, mutazioni proteiche