Clear Sky Science · en
G-protein activation of the dark-state conformation of the visual G protein-coupled receptor rhodopsin by releasing critical structural constraints
Why a Night-Vision Protein Matters
Seeing in near darkness depends on a light-sensing protein in our eyes called rhodopsin. Normally, rhodopsin stays almost completely silent until a photon of light hits it, helping us avoid visual “noise” that would blur dim scenes. But in certain inherited eye diseases, rhodopsin becomes too active even in the dark, confusing the visual system and impairing night vision. This study dissects how tiny changes in the structure of rhodopsin can flip it from a tightly locked, quiet state into an active, signaling state—without any light at all—offering new insight into how our vision system keeps its extraordinary sensitivity under control. 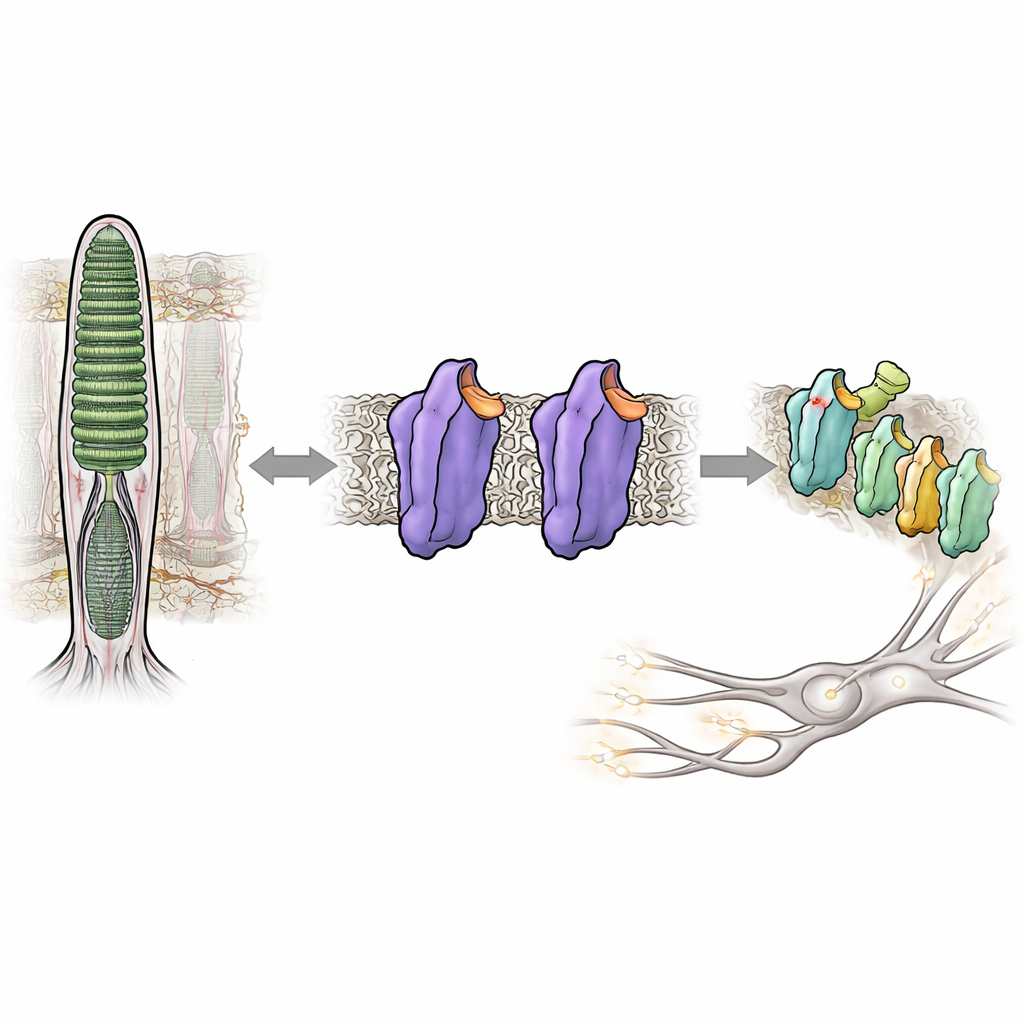
A Molecular Light Switch in the Eye
Rhodopsin belongs to a huge family of cellular switches called G protein–coupled receptors, which respond to hormones, odors, and many drugs. In rod cells of the retina, rhodopsin is tuned to respond to the faintest glimmer of light. It does this by binding a small vitamin A–derived molecule, 11-cis-retinal, that acts as a built-in brake, holding the protein in a quiet, or dark, state. When light hits, the retinal snaps into a new shape, triggering a cascade of structural changes in rhodopsin and turning it into an active form known as Meta II. This active form then switches on a partner protein, a G protein called transducin, starting the electrical signal that the brain interprets as vision.
When the Dark State Loosens
Some people inherit small changes, or mutations, in rhodopsin that weaken this dark brake and let the protein signal too much even without light. Such “leaky” activity is linked to disorders like congenital stationary night blindness and retinitis pigmentosa. The authors focused on three positions within rhodopsin that function as internal “microswitches,” each helping hold the protein in its inactive shape. Individually, known mutations at these sites can mildly destabilize the dark state or increase spontaneous activity. Here, the researchers combined them into engineered double and triple mutants to see whether simultaneously relaxing several constraints would push rhodopsin into an active-like state entirely in the dark.
Building a Permanently Primed Rhodopsin
The team produced mutant rhodopsins in cultured cells, purified them, and reassembled them with their normal retinal cofactor. Using ultraviolet–visible spectroscopy, they tracked how these pigments absorbed light, how stable they were at higher temperatures, and how they responded to light exposure and acidic conditions. The triple mutant, carrying all three substitutions, showed a dominant absorption band matching that of the active Meta II form, even without illumination. It was thermally unstable, quickly losing its dark-state spectral signature, consistent with a protein that readily shifts toward an active shape. Remarkably, this mutant could even bind all-trans-retinal—the light-activated form of the chromophore—in the dark, something normal rhodopsin cannot do, further indicating an already “open” conformation. Functional tests confirmed that this triple mutant fully activated the G protein in the dark and performed as well as or better than normal rhodopsin after light exposure. 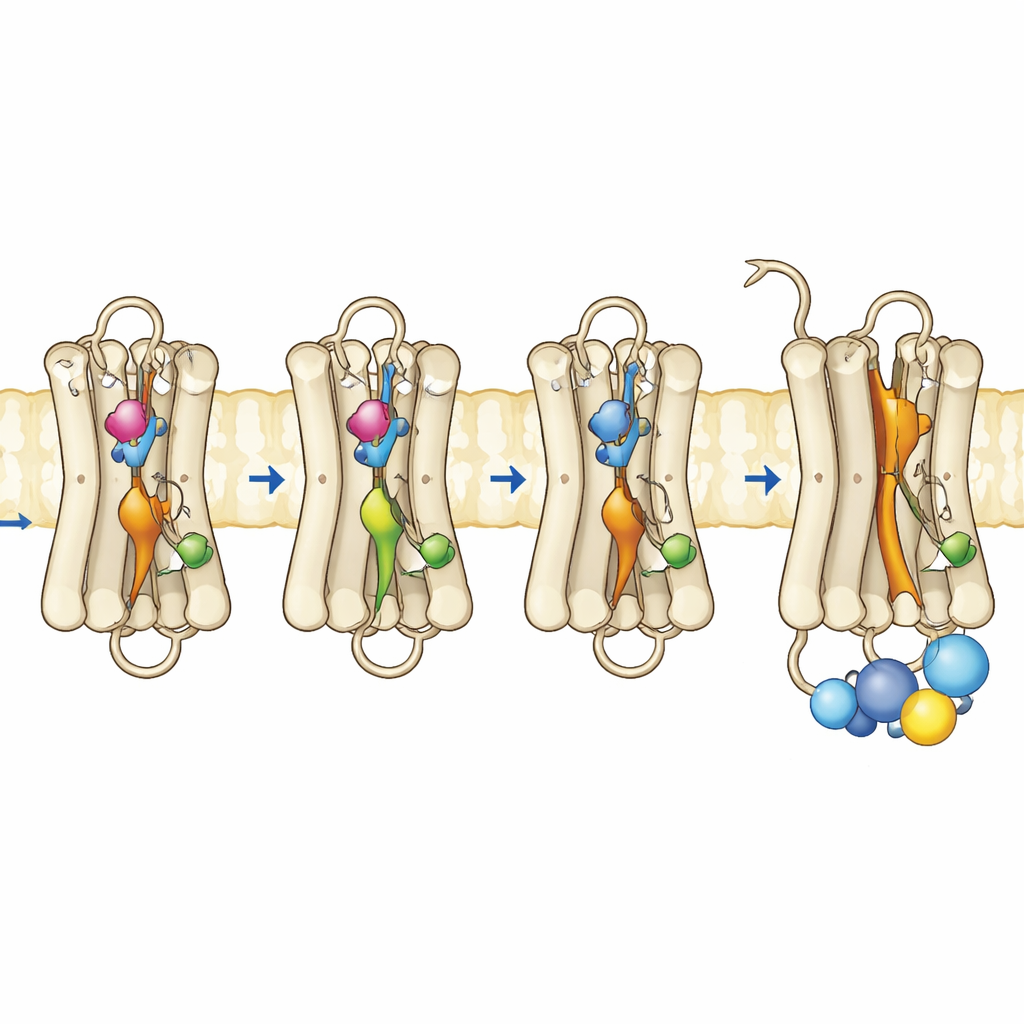
Watching the Protein Move
To see what changed at the level of motion and flexibility, the authors used fluorescence-based methods that report on how a specific helix near the inner surface of rhodopsin, called helix 8, moves and how a fluorescent peptide fragment of the G protein behaves when it approaches the receptor. In normal rhodopsin, strong fluorescence changes appear only after light turns on the receptor. In contrast, the triple mutant and one of the single-site mutants already showed active-like behavior in the dark, indicating that the inner surface where the G protein docks had reorganized. Time-resolved fluorescence anisotropy measurements revealed that helix 8 in the triple mutant was more mobile and occupied a different conformational space, resembling that of active rhodopsin. Complementary computer simulations of the protein in a membrane supported these findings: the three mutations cooperatively disrupted key internal contacts, allowing several internal switches to adopt active-like geometries even without a light-triggered retinal flip.
What This Means for Vision and Disease
Together, the spectroscopic measurements, functional assays, fluorescence studies, and simulations show that altering just three carefully chosen sites is enough to unlock rhodopsin’s dark state and drive it into an active-like conformation on its own. In essence, these mutations relax critical internal braces that normally keep the receptor quiet until light arrives. This work clarifies how only a few residues among more than 200 amino acids can control the transition between silence and signaling in a key visual protein. Understanding these structural levers helps explain how certain inherited mutations cause excessive activity and night-vision problems, and it offers a more general blueprint for how G protein–coupled receptors balance stability with responsiveness throughout the body.
Citation: Ramon, E., Kirchberg, K., Jiménez-Rosés, M. et al. G-protein activation of the dark-state conformation of the visual G protein-coupled receptor rhodopsin by releasing critical structural constraints. Commun Biol 9, 523 (2026). https://doi.org/10.1038/s42003-026-09774-w
Keywords: rhodopsin, night vision, G protein–coupled receptors, retinal diseases, protein mutations