Clear Sky Science · sv
G-proteinaktivering av mörkertillståndet hos den visuella G-proteinkopplade receptorn rodopsin genom att frigöra kritiska strukturella begränsningar
Varför ett nattseendeprotein är viktigt
Att se i nästan mörker beror på ett ljuskänsligt protein i våra ögon som kallas rodopsin. Normalt förblir rodopsin nästan helt tyst tills en foton träffar det, vilket hjälper oss undvika visuellt ”brus” som skulle sudda ut dunkla scener. Men vid vissa ärftliga ögonsjukdomar blir rodopsin överaktivt även i mörker, vilket förvirrar synsystemet och försämrar nattseendet. Denna studie dissekerar hur små förändringar i rodopsins struktur kan växla det från ett tätt låst, tyst tillstånd till ett aktivt, signalerande tillstånd—utan något ljus alls—och ger ny insikt i hur vårt visuella system håller sin extraordinära känslighet under kontroll. 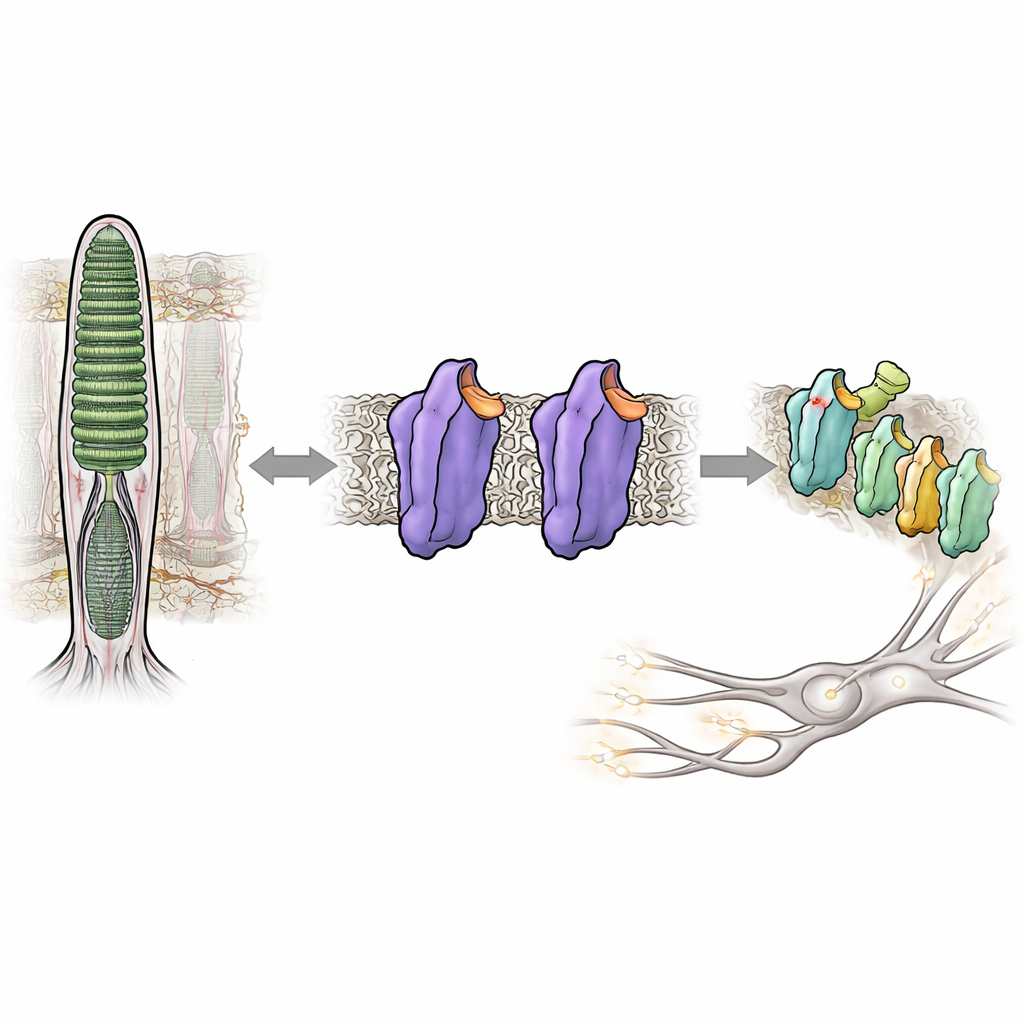
En molekylär ljusbrytare i ögat
Rodopsin tillhör en stor familj av cellulära brytare som kallas G-proteinkopplade receptorer, vilka svarar på hormoner, dofter och många läkemedel. I stavcellerna i näthinnan är rodopsin finjusterad för att reagera på den svagaste ljusglimten. Det gör det genom att binda en liten vitamin A‑ härledd molekyl, 11-cis-retinal, som fungerar som en inbyggd broms och håller proteinet i ett tyst eller mörkertillstånd. När ljus träffar, slår retinalen om till en ny form och utlöser en kaskad av strukturella förändringar i rodopsin och omvandlar det till en aktiv form känd som Meta II. Denna aktiva form slår sedan på en partnerprotein, ett G-protein kallat transducin, vilket startar den elektriska signal som hjärnan tolkar som syn.
När mörkertillståndet lossnar
Vissa personer ärver små förändringar, eller mutationer, i rodopsin som försvagar denna mörkerbroms och låter proteinet signalera för mycket även utan ljus. Sådan ”läckande” aktivitet kopplas till störningar som kongenital stationär nattblindhet och retinitis pigmentosa. Författarna fokuserade på tre positioner inom rodopsin som fungerar som interna ”mikrobrytare”, var och en hjälper till att hålla proteinet i dess inaktiva form. Enskilda, kända mutationer på dessa platser kan mildra mörkertillståndets stabilitet eller öka spontan aktivitet. Här kombinerade forskarna dem till konstruerade dubbel- och trippelmutanter för att se om samtidig avslappning av flera begränsningar skulle pressa rodopsin in i ett aktivliknande tillstånd helt i mörker.
Att bygga en permanent förberedd rodopsin
Teamet producerade mutanta rodopsiner i odlingsceller, renade dem och återmonterade dem med deras normala retinal-ko-faktor. Med hjälp av ultraviolett–visibel spektroskopi följde de hur dessa pigment absorberade ljus, hur stabila de var vid högre temperaturer och hur de svarade på ljusexponering och sura förhållanden. Trippelmutanten, med alla tre substitutioner, visade ett dominerande absorptionsband som matchade det aktiva Meta II-formens, även utan belysning. Den var termiskt instabil och förlorade snabbt sitt mörkertillstånds spektrala signatur, vilket stämmer överens med ett protein som lätt skiftar mot en aktiv form. Anmärkningsvärt nog kunde denna mutant till och med binda all-trans-retinal—den ljusaktiverade formen av kromoforen—i mörker, något normalt rodopsin inte kan göra, vilket ytterligare indikerar en redan ”öppen” konformation. Funktionella tester bekräftade att denna trippelmutant helt aktiverade G-proteinet i mörker och presterade lika bra eller bättre än normalt rodopsin efter ljusexponering. 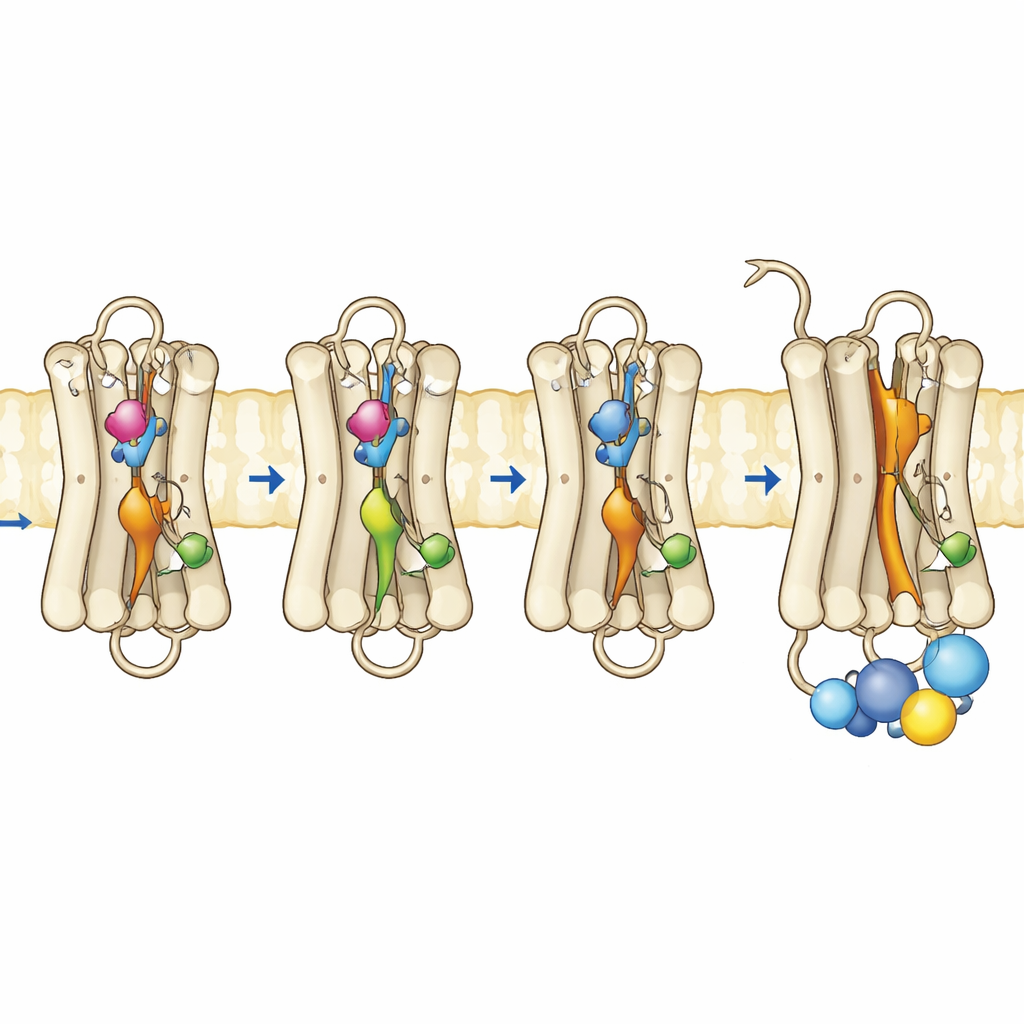
Att se proteinet röra sig
För att se vad som förändrats på nivån av rörelse och flexibilitet använde författarna fluorescensbaserade metoder som rapporterar hur en specifik helix nära rodopsins inre yta, kallad helix 8, rör sig och hur en fluorescerande peptidbit från G-proteinet beter sig när den närmar sig receptorn. I normalt rodopsin uppstår starka fluorescensförändringar först efter att ljus aktiverat receptorn. I kontrast visade trippelmutanten och en av enkelplatsmutanterna redan aktivliknande beteende i mörker, vilket indikerar att den inre ytan där G-proteinet dockar hade omorganiserats. Tidsupplösta mätningar av fluorescensanisotropi avslöjade att helix 8 i trippelmutanten var mer rörlig och upptog ett annat konformationsutrymme, liknande det hos aktivt rodopsin. Kompletterande datorsimuleringar av proteinet i ett membran stödde dessa fynd: de tre mutationerna störde kooperativt viktiga interna kontakter och tillät flera interna brytare anta aktivliknande geometrier även utan en ljusutlöst retinalomslagning.
Vad detta betyder för syn och sjukdom
Tillsammans visar de spektroskopiska mätningarna, funktionella analyserna, fluorescensstudierna och simuleringarna att ändring av bara tre noggrant utvalda platser är tillräckligt för att låsa upp rodopsins mörkertillstånd och driva det in i en aktivliknande konformation på egen hand. I korthet slappnar dessa mutationer av kritiska inre stag som normalt håller receptorn tyst tills ljus anländer. Detta arbete klargör hur bara några få restpositions bland mer än 200 aminosyror kan kontrollera övergången mellan tystnad och signalering i ett nyckelprotein för synen. Att förstå dessa strukturella hävstänger hjälper till att förklara hur vissa ärftliga mutationer orsakar överdriven aktivitet och problem med nattseendet, och ger en mer generell plan för hur G-proteinkopplade receptorer balanserar stabilitet och lyhördhet i hela kroppen.
Citering: Ramon, E., Kirchberg, K., Jiménez-Rosés, M. et al. G-protein activation of the dark-state conformation of the visual G protein-coupled receptor rhodopsin by releasing critical structural constraints. Commun Biol 9, 523 (2026). https://doi.org/10.1038/s42003-026-09774-w
Nyckelord: rodopsin, nattseende, G-proteinkopplade receptorer, retinala sjukdomar, proteinmutationer