Clear Sky Science · fr
Activation par la protéine G de la conformation en état sombre du récepteur visuel couplé aux protéines G rhodopsine en levant des contraintes structurelles critiques
Pourquoi une protéine de vision nocturne compte
La capacité à voir dans presque l’obscurité dépend d’une protéine photodétectrice de nos yeux appelée rhodopsine. Normalement, la rhodopsine reste presque complètement silencieuse jusqu’à ce qu’un photon la frappe, ce qui nous aide à éviter le « bruit » visuel qui brouillerait les scènes faiblement éclairées. Mais dans certaines maladies héréditaires de l’œil, la rhodopsine devient trop active même dans l’obscurité, perturbant le système visuel et altérant la vision nocturne. Cette étude dissèque comment de minuscules modifications de la structure de la rhodopsine peuvent la faire basculer d’un état verrouillé et silencieux vers un état actif et signalant — sans aucune lumière — offrant un nouvel éclairage sur la manière dont notre système visuel maintient sa sensibilité exceptionnelle sous contrôle. 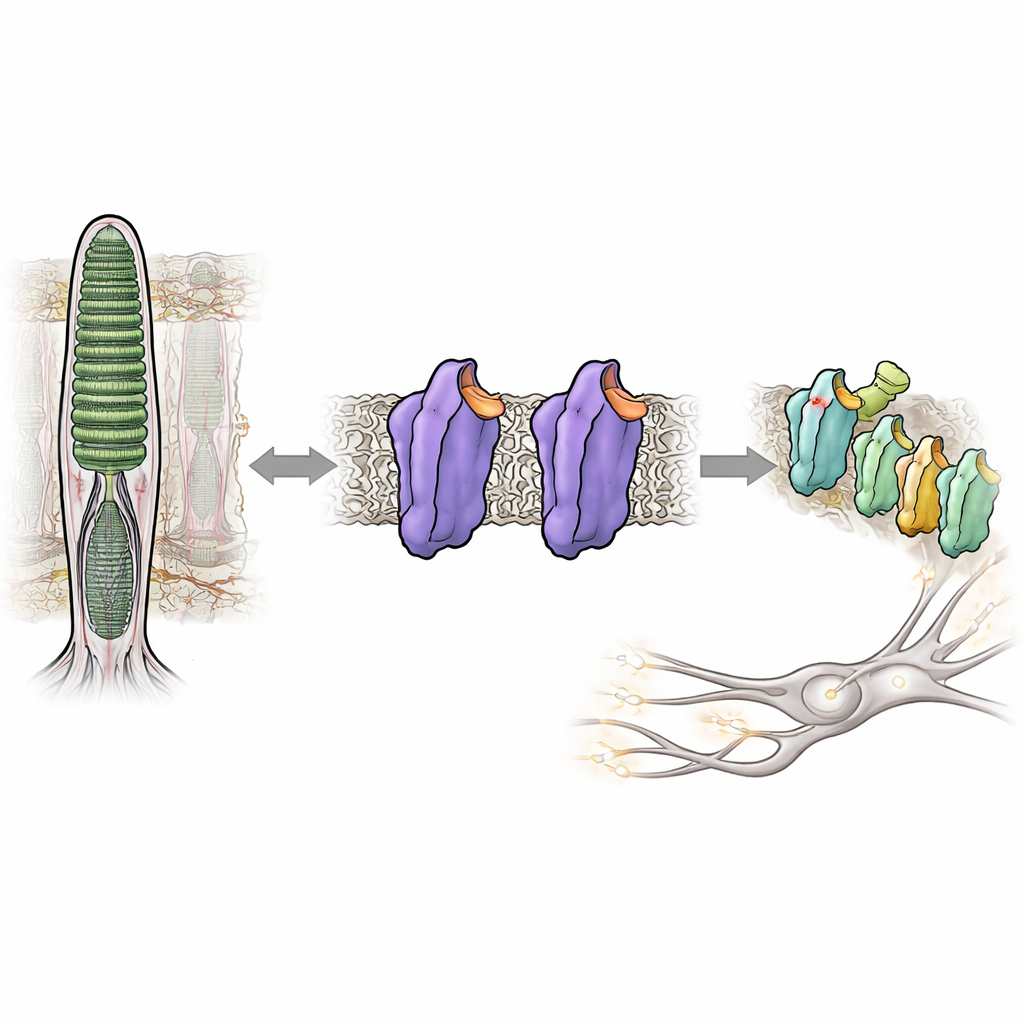
Un interrupteur moléculaire dans l’œil
La rhodopsine appartient à une vaste famille d’interrupteurs cellulaires appelés récepteurs couplés aux protéines G, qui répondent aux hormones, aux odeurs et à de nombreux médicaments. Dans les bâtonnets de la rétine, la rhodopsine est réglée pour répondre au moindre scintillement de lumière. Elle y parvient en liant une petite molécule dérivée de la vitamine A, le 11-cis-rétinal, qui agit comme un frein interne, maintenant la protéine dans un état calme, ou état sombre. Quand la lumière frappe, le rétinal bascule en une nouvelle conformation, déclenchant une cascade de changements structuraux dans la rhodopsine et la convertissant en une forme active connue sous le nom de Meta II. Cette forme active active ensuite une protéine partenaire, une protéine G appelée transducine, amorçant le signal électrique que le cerveau interprète comme la vision.
Quand l’état sombre se relâche
Certaines personnes héritent de petites modifications, ou mutations, dans la rhodopsine qui affaiblissent ce frein sombre et laissent la protéine signaler excessivement même sans lumière. Une activité « qui fuit » est liée à des troubles comme la cécité nocturne stationnaire congénitale et la rétinite pigmentaire. Les auteurs se sont concentrés sur trois positions au sein de la rhodopsine qui fonctionnent comme des « microswitchs » internes, chacun contribuant à maintenir la protéine dans sa forme inactive. Pris individuellement, des mutations connues en ces sites peuvent légèrement déstabiliser l’état sombre ou augmenter l’activité spontanée. Ici, les chercheurs les ont combinées en mutants doubles et triples conçus pour tester si la détente simultanée de plusieurs contraintes pousserait la rhodopsine vers un état de type actif entièrement dans le noir.
Construire une rhodopsine en permanence armée
L’équipe a produit des rhodopsines mutantes dans des cellules en culture, les a purifiées et les a réassemblées avec leur cofacteur rétinien normal. En utilisant la spectroscopie ultraviolet–visible, ils ont suivi comment ces pigments absorbaient la lumière, leur stabilité à des températures élevées et leur réponse à l’exposition lumineuse et aux conditions acides. Le mutant triple, portant les trois substitutions, a présenté une bande d’absorption dominante correspondant à celle de la forme active Meta II, et ce sans illumination. Il était thermiquement instable, perdant rapidement sa signature spectrale de l’état sombre, ce qui correspond à une protéine qui bascule facilement vers une conformation active. De façon remarquable, ce mutant pouvait même lier le tout-trans-rétinal — la forme du chromophore activée par la lumière — dans l’obscurité, ce que la rhodopsine normale ne peut pas faire, indiquant encore une conformation déjà « ouverte ». Des tests fonctionnels ont confirmé que ce mutant triple activait pleinement la protéine G dans l’obscurité et fonctionnait aussi bien, voire mieux, que la rhodopsine normale après exposition à la lumière. 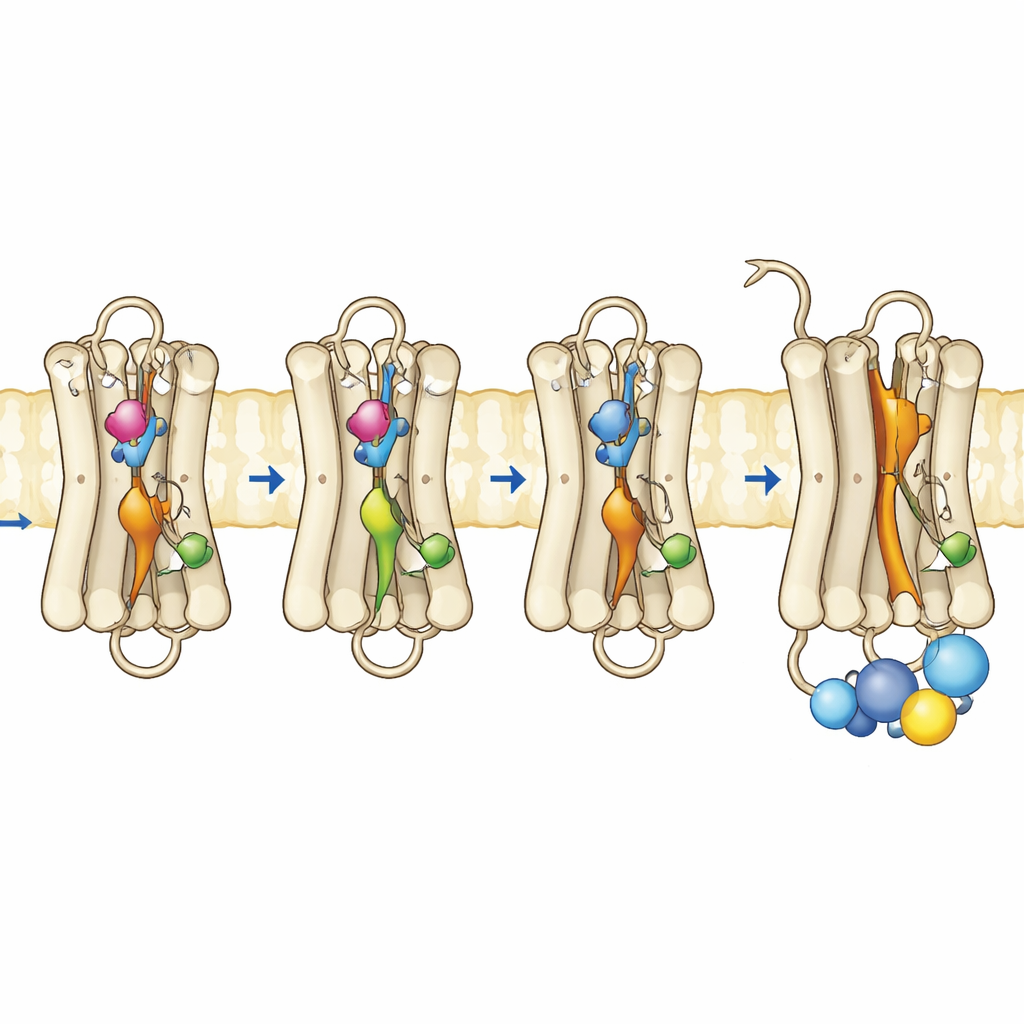
Observer le mouvement de la protéine
Pour voir ce qui changeait au niveau du mouvement et de la flexibilité, les auteurs ont utilisé des méthodes basées sur la fluorescence qui renseignent sur la façon dont une hélice spécifique près de la surface interne de la rhodopsine, appelée hélice 8, se déplace et sur le comportement d’un fragment peptidique fluorescent de la protéine G lorsqu’il s’approche du récepteur. Dans la rhodopsine normale, de fortes variations de fluorescence n’apparaissent qu’après que la lumière a activé le récepteur. En revanche, le mutant triple et l’un des mutants simples présentaient déjà en obscurité un comportement de type actif, indiquant que la surface interne où la protéine G s’amarrage s’était réorganisée. Des mesures d’anisotropie de fluorescence résolues dans le temps ont révélé que l’hélice 8 du mutant triple était plus mobile et occupait un espace conformationnel différent, ressemblant à celui de la rhodopsine active. Des simulations informatiques complémentaires de la protéine dans une membrane ont étayé ces observations : les trois mutations perturbaient de manière coopérative des contacts internes clés, permettant à plusieurs microswitchs internes d’adopter des géométries de type actif même sans basculement du rétinal déclenché par la lumière.
Ce que cela signifie pour la vision et la maladie
Ensemble, les mesures spectroscopiques, les essais fonctionnels, les études de fluorescence et les simulations montrent que modifier seulement trois sites choisis avec soin suffit à déverrouiller l’état sombre de la rhodopsine et à la pousser dans une conformation de type actif par elle-même. En substance, ces mutations relâchent des contreforts internes critiques qui, normalement, gardent le récepteur silencieux jusqu’à l’arrivée de la lumière. Ce travail clarifie comment quelques résidus, au sein de plus de 200 acides aminés, peuvent contrôler la transition entre silence et signalisation dans une protéine visuelle clé. Comprendre ces leviers structurels aide à expliquer comment certaines mutations héréditaires provoquent une activité excessive et des problèmes de vision nocturne, et offre un modèle plus général de la façon dont les récepteurs couplés aux protéines G équilibrent stabilité et réactivité dans tout l’organisme.
Citation: Ramon, E., Kirchberg, K., Jiménez-Rosés, M. et al. G-protein activation of the dark-state conformation of the visual G protein-coupled receptor rhodopsin by releasing critical structural constraints. Commun Biol 9, 523 (2026). https://doi.org/10.1038/s42003-026-09774-w
Mots-clés: rhodopsine, vision nocturne, récepteurs couplés aux protéines G, maladies rétiniennes, mutations protéiques