Clear Sky Science · ru
Активация Г-белком темновой конформации визуального рецептора, связанного с G-белком, родопсина путем снятия критических структурных ограничений
Почему белок ночного зрения важен
Возможность видеть в полумраке зависит от светочувствительного белка в наших глазах — родопсина. В норме родопсин почти полностью молчит до тех пор, пока не попадет фотон света, что помогает избегать визуального «шума», размывающего темные сцены. Но при некоторых наследственных заболеваниях глаз родопсин становится чрезмерно активным даже в темноте, дезорганизуя зрительную систему и ухудшая ночное зрение. В этом исследовании показано, как крошечные изменения в структуре родопсина могут перевести его из плотно зафиксированного, тихого состояния в активную сигнальную конформацию — без какого-либо света — что дает новое понимание того, как наша зрительная система поддерживает свою исключительную чувствительность под контролем. 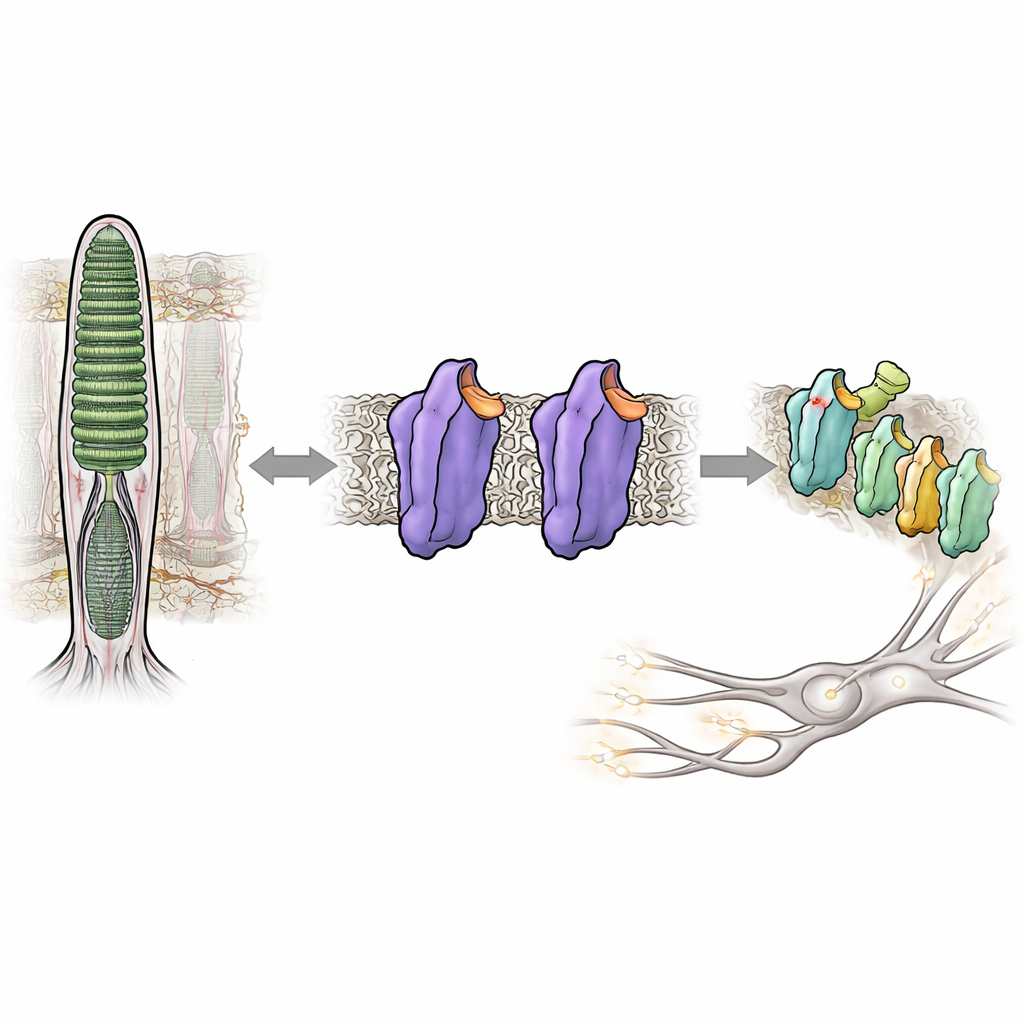
Молекулярный световой переключатель в глазу
Родопсин принадлежит к огромному семейству клеточных переключателей, называемых рецепторами, связанными с G-белком, которые реагируют на гормоны, запахи и многие лекарства. В палочках сетчатки родопсин настроен реагировать на самый слабый отблеск света. Это достигается за счет связывания с небольшим производным витамина A, 11-цис-ретиналем, который действует как встроенный тормоз, удерживающий белок в тихой, или темновой, конформации. Когда попадает свет, ретинал мгновенно меняет форму, вызывая каскад структурных перестроек в родопсине и переходя его в активную форму, известную как Мета II. Эта активная форма затем включает партнерский белок — G-белок трандуксин, запускающий электрический сигнал, который мозг интерпретирует как зрение.
Когда темновое состояние ослабляется
Некоторые люди наследуют небольшие изменения, или мутации, в родопсине, которые ослабляют этот темновой тормоз и позволяют белку избыточно сигнализировать даже без света. Такая «текучая» активность связана с расстройствами, такими как врожденная стационарная ночная слепота и ретинит пигментоза. Авторы сосредоточились на трех позициях в родопсине, которые функционируют как внутренние «микропереключатели», каждая из которых помогает удерживать белок в неактивной форме. По отдельности известные мутации в этих участках могут слегка дестабилизировать темновое состояние или увеличивать спонтанную активность. В этой работе исследователи совместили их в конструируемых двойных и тройных мутантах, чтобы проверить, приведет ли одновременное ослабление нескольких ограничений к переходу родопсина в активоподобное состояние полностью в темноте.
Создание постоянно подготовленного родопсина
Команда получила мутантные родопсины в культивируемых клетках, очистила их и заново собрала с их нормальным ретинальным кофактором. С помощью ультрафиолетово-видимой спектроскопии они отслеживали, как эти пигменты поглощают свет, насколько они устойчивы при повышенных температурах и как реагируют на воздействие света и кислую среду. Тройной мутант, несущий все три замены, показал доминирующую полосу поглощения, соответствующую активной форме Мета II, даже без освещения. Он был термально нестабилен, быстро теряя спектральную подпись темнового состояния, что согласуется с тем, что белок охотно смещается в активную форму. Удивительно, но этот мутант даже мог связывать all-trans-ретиналь — световоактивную форму хромофора — в темноте, чего нормальный родопсин сделать не может, что дополнительно указывает на уже «открытую» конформацию. Функциональные тесты подтвердили, что этот тройной мутант полностью активировал G-белок в темноте и работал не хуже или лучше, чем нормальный родопсин после воздействия света. 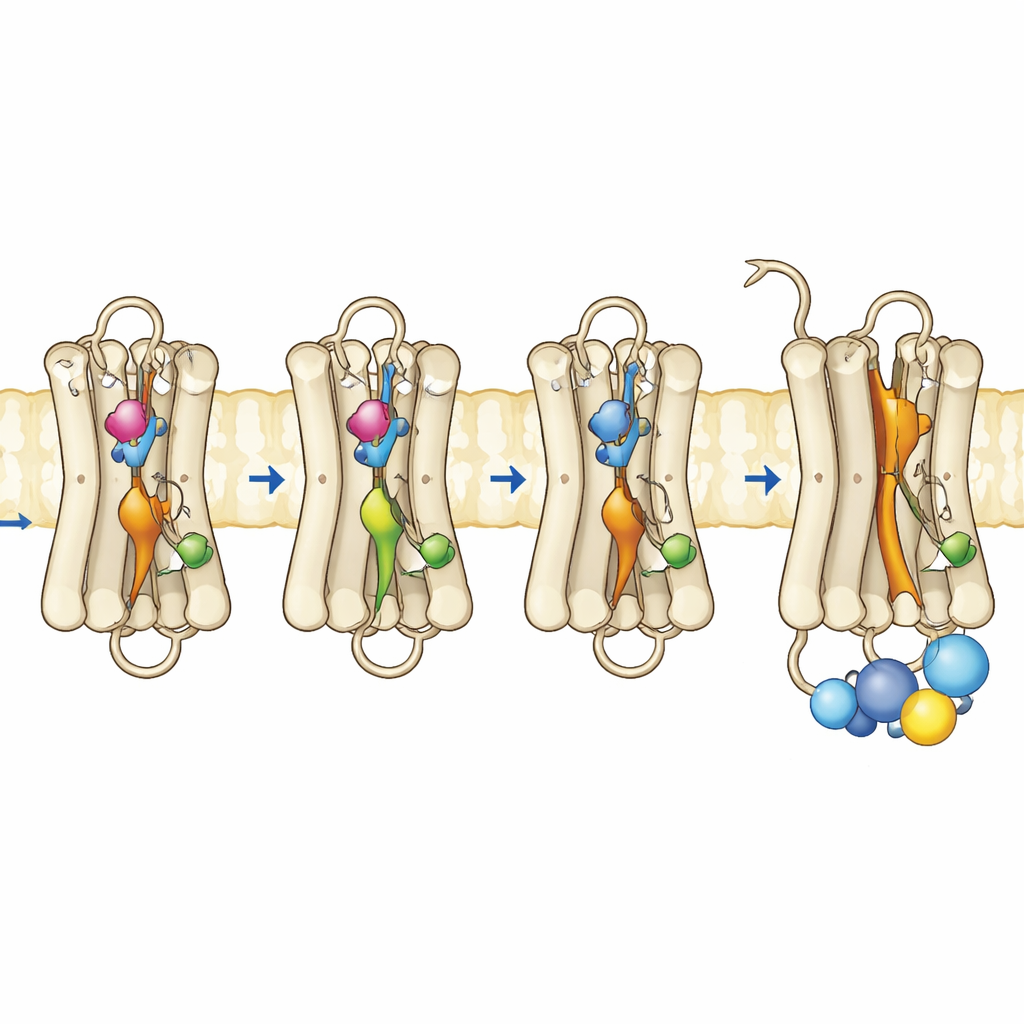
Наблюдая за движением белка
Чтобы увидеть, что изменилось на уровне движений и гибкости, авторы использовали флуоресцентные методы, которые показывают, как движется конкретная спираль у внутренней поверхности родопсина, называемая спиралью 8, и как ведет себя флуоресцентный пептидный фрагмент G-белка при приближении к рецептору. В нормальном родопсине резкие изменения флуоресценции появляются только после включения рецептора светом. Напротив, тройной мутант и один из одноточечных мутантов уже демонстрировали активоподобное поведение в темноте, что указывает на реорганизацию внутренней поверхности, где докируется G-белок. Временнo-разрешенная измерения флуоресцентной анизотропии показали, что спираль 8 в тройном мутанте была более подвижна и занимала другую конформационную область, напоминающую активный родопсин. Дополняющие компьютерные симуляции белка в мембране подтвердили эти наблюдения: три мутации кооперативно нарушали ключевые внутренние контакты, позволяя нескольким внутренним переключателям принять активоподобные геометрии даже без светового переворота ретиналя.
Что это означает для зрения и болезней
В совокупности спектроскопические измерения, функциональные анализы, флуоресцентные исследования и симуляции показывают, что изменение всего трех тщательно подобранных участков достаточно, чтобы разблокировать темповое состояние родопсина и самостоятельно перевести его в активоподобную конформацию. По сути, эти мутации ослабляют критические внутренние распорки, которые обычно удерживают рецептор в спокойствии до прихода света. Эта работа проясняет, как всего несколько остатков среди более чем 200 аминокислот могут контролировать переход между молчанием и сигнализацией в ключевом зрительном белке. Понимание этих структурных рычагов помогает объяснить, почему некоторые наследственные мутации вызывают избыточную активность и проблемы с ночным зрением, а также дает более общую схему того, как рецепторы, связанные с G-белком, уравновешивают стабильность и отзывчивость по всему организму.
Цитирование: Ramon, E., Kirchberg, K., Jiménez-Rosés, M. et al. G-protein activation of the dark-state conformation of the visual G protein-coupled receptor rhodopsin by releasing critical structural constraints. Commun Biol 9, 523 (2026). https://doi.org/10.1038/s42003-026-09774-w
Ключевые слова: родопсин, ночное зрение, рецепторы, связанные с G-белком, заболевания сетчатки, мутации белков