Clear Sky Science · zh
通过选择性序列实现证据累积的神经回路模型
大脑如何悄然累加线索
从过马路到选零食,日常决策都依赖于大脑在一段时间内悄然将各类线索相加的能力。数十年来,神经科学家普遍认为这是由一群持续活跃的神经元完成的,像一个缓慢上升的心理仪表指向决策。然而,本研究提出了当这一经典图景失效时会发生什么——当脑细胞以快速、不断变化的模式放电时——并揭示了大脑可能用来在证据展开时持续跟踪信息的新型回路设计。
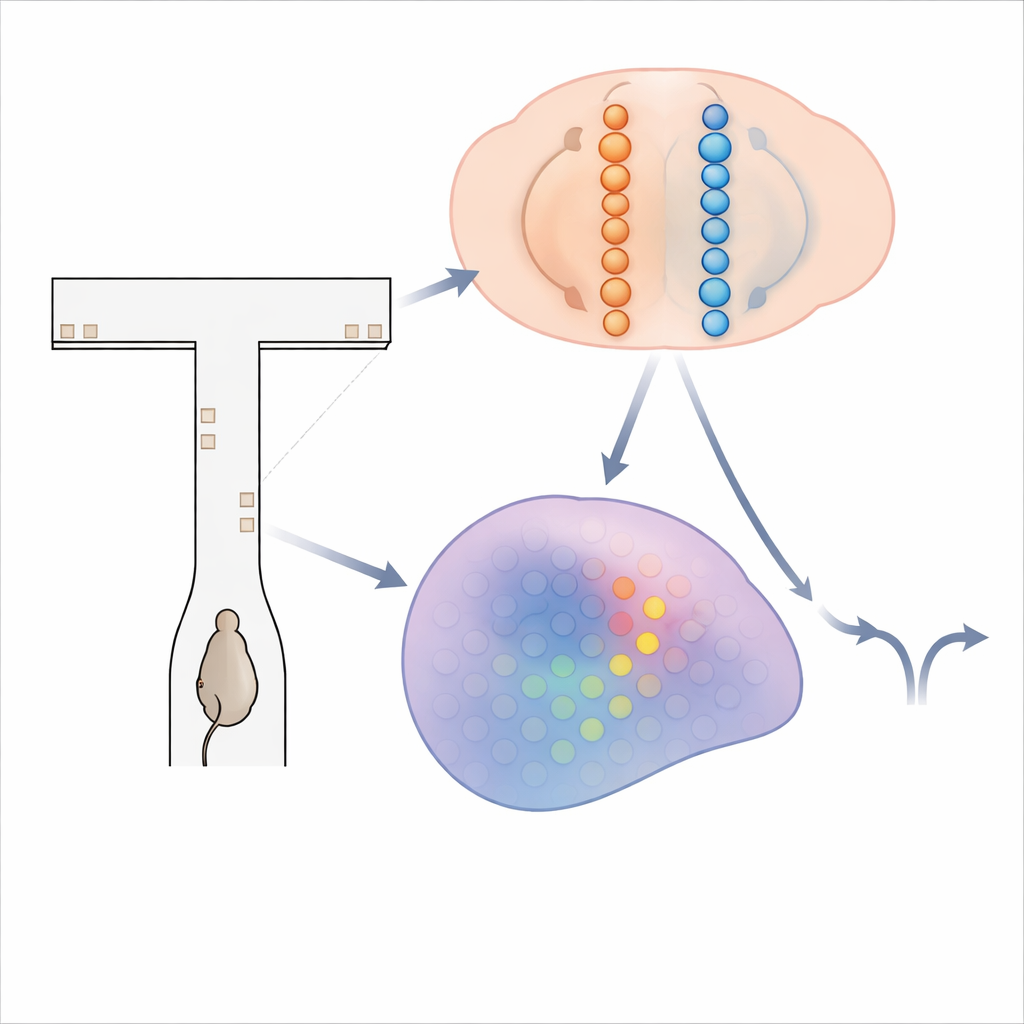
闪烁线索的迷宫
为探查大脑如何累积证据,研究者让小鼠在一个虚拟的T形迷宫中奔跑。随着小鼠沿迷宫的长茎前进,左右墙上会短暂出现视觉“塔状”刺激。动物的目标很简单:到达尽头时,朝出现塔较多的一侧转向以获得奖励。要完成这个任务,需在数秒内将出现的塔相加,并在最终转向前的延迟期中维持这个累积的计数。在训练有素的小鼠完成数百次试次的同时,团队使用先进的钙成像技术记录了来自四个与决策和记忆相关脑区的超过14,000个神经元的活动。
从平滑斜坡到竞速波动
受猴子和其他动物记录启发的经典理论提出,单个神经元的发放率会随着证据朝某一选择累积而逐渐上升或下降,呈现缓慢的斜坡状。但在这些小鼠数据中,这种解释并不成立。皮层、海马和纹状体的神经元并非持续缓慢上升,而是倾向于短暂依次放电,形成与动物沿迷宫行进及其最终选择相关的有序序列。某个细胞可能在走廊中段为随后会左转的小鼠发放,而另一个细胞则在更后面为右转发放。这些“选择性序列”随试次推进在各脑区扫过,挑战了由单一稳定神经群维持决策变量的观念。
两种实现累加的回路蓝图
为解释变化的细胞序列如何仍能携带精确的累积证据,作者建立并分析了两类神经回路模型。第一类称为“竞争链条”,证据存储在两条并行神经链中活动强度的相对差异上,一条偏向左、一条偏向右。一个与位置相关的门控信号确保在迷宫的每个位置仅有一对小神经元(每条链各一个)被激活。进入的塔状刺激会将这对活跃神经元朝某一侧推动,特化的连接把这种累积的不平衡传递给动物前进时的下一对神经元。在这种设计中,证据量以单调方式编码:越多的右侧证据只是意味着右链活动更强、左链更弱。第二种模型为“位置门控峰”,迷宫的每个位置对应一张神经元“薄片”,这些神经元沿一个证据轴共同形成一个紧凑的活动峰。塔刺激使活动峰向左或向右移动,像在刻度上滑动一个标记。随着小鼠前进,前馈连接将该活动峰传递给下一张薄片,保持其位置。在这里,某一时刻哪些特定神经元活跃直接表示证据水平,产生窄且峰状的调谐,而不是宽广的斜坡。
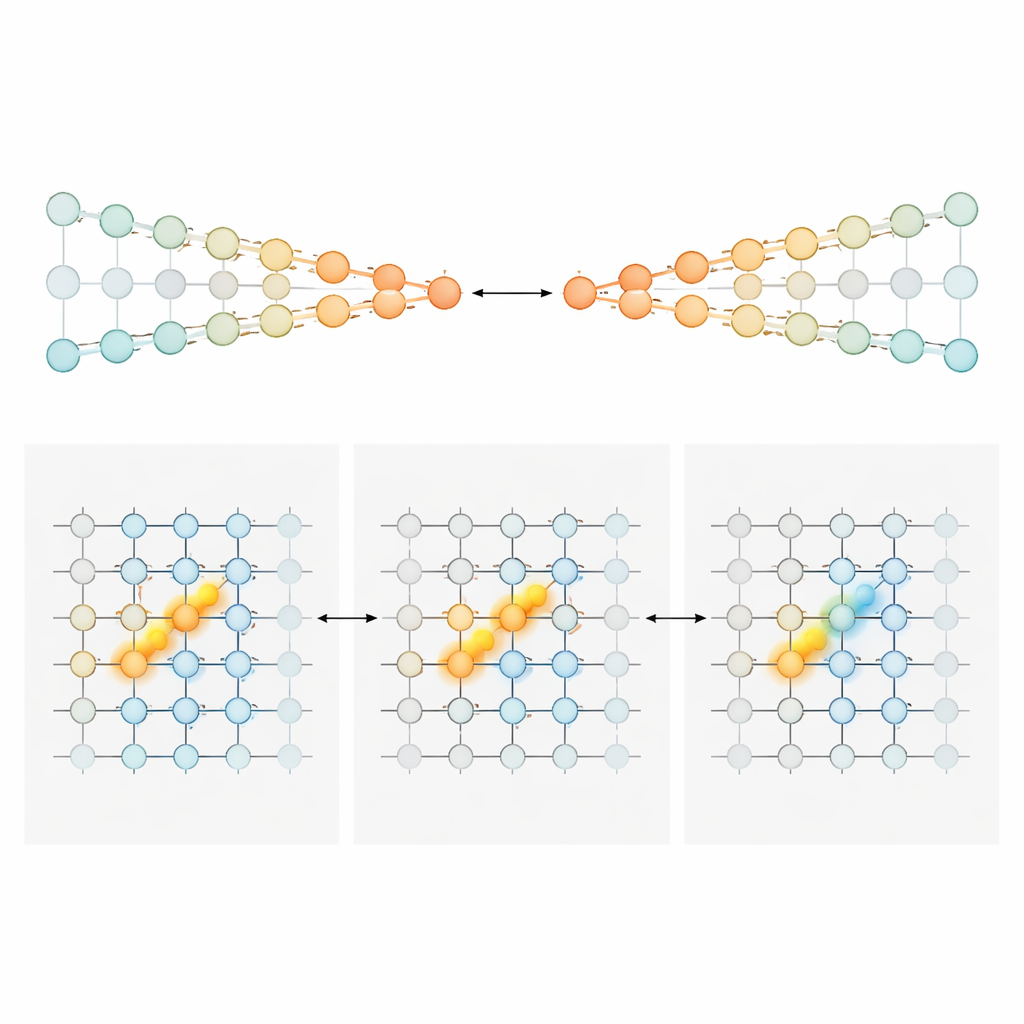
不同脑区,不同计数风格
有了这些蓝图,团队回过头检查记录数据以判断各脑区更像哪种模型。在前扣带皮层和后扣带皮层——类似额叶与顶叶、参与规划与导航的区域——大多数神经元表现出较为宽泛且主要单向的证据敏感性。它们的活动倾向随着对偏好选择支持度的增强而稳步上升,与竞争链条式的编码相符。在提示期的早期,这些群体携带关于“更多多少”偏向一侧的渐进信号;当小鼠接近选择点时,群体反应变得更尖锐,逐渐塌缩为更具类别性的、类似选择的信号。截然不同的是,海马神经元通常对特定证据值呈现窄的钟形响应,不同细胞覆盖整个证据范围。这一模式正好符合峰模型的预测,也与海马在绘制动物位置与路径上发生事件的更广泛角色一致。在延迟期主要被记录的纹状体神经元则倾向于以更全或无式的方式与最终转向相关发放,符合其在动作选择中作为下游角色的说法。
对理解决策的意义
该研究的核心信息是,大脑可能在同一任务中使用多种回路技巧来跟踪展开的证据。某些区域似乎将证据表示为竞争神经池之间的平滑博弈,适合比较选项并驱动最终选择;另一些区域,如海马,则以更像地图的方式表示证据,活跃细胞的身份直接指示当前的计数。两种方案都依赖于瞬时活动的序列而非静态放电,且通过精心构造的连接在任务的内部“时钟”前进时传递信息。结合这些模型与测量结果,表明我们的决策源于灵活的、具区域特异性的机制,既能将嘈杂的线索相加,也能在恰当的时间将累积结果路由到恰当的位置。
引用: Brown, L.S., Cho, J.R., Bolkan, S.S. et al. Neural circuit models for evidence accumulation through choice-selective sequences. Nat Commun 17, 4055 (2026). https://doi.org/10.1038/s41467-026-70267-9
关键词: 决策, 证据累积, 神经回路, 海马体, 皮层序列