Clear Sky Science · it
Modelli di circuiti neurali per l’accumulazione di prove tramite sequenze selettive per la scelta
Come il cervello conta silenziosamente gli indizi
Scelte quotidiane, dal attraversare una strada allo scegliere uno snack, dipendono dalla capacità del cervello di sommare silenziosamente frammenti di evidenza nel tempo. Per decenni, i neuroscienziati hanno pensato che questo fosse fatto da gruppi di neuroni che restavano attivi in modo stabile, come un indicatore mentale che lentamente sale verso una decisione. Questo studio chiede cosa succede quando quella visione classica è sbagliata—quando le cellule cerebrali invece sparano in schemi rapidi e mutevoli—e rivela nuovi progetti di circuito che il cervello potrebbe usare per tenere traccia delle evidenze mentre si dispiegano.
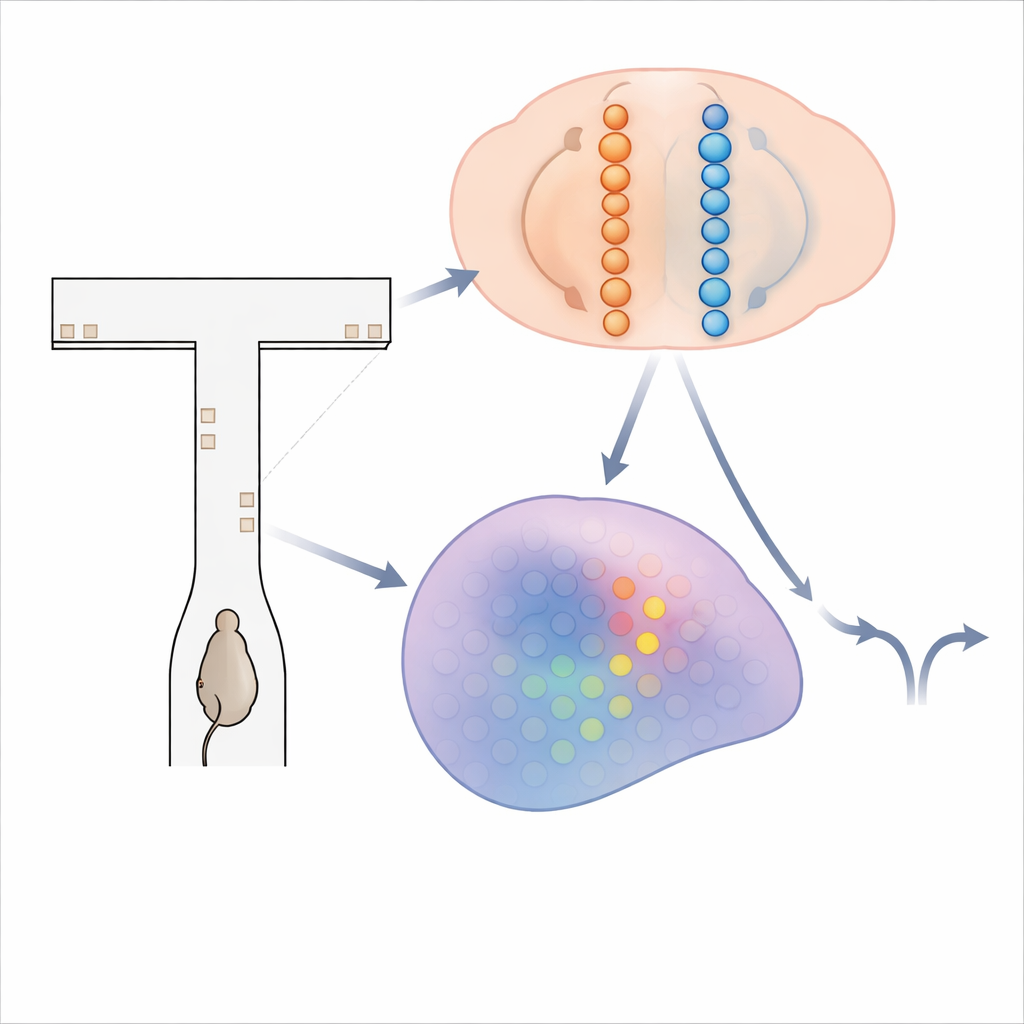
Un labirinto di indizi che sfarfallano
Per indagare come il cervello accumula evidenze, i ricercatori hanno usato topi che correvano attraverso un labirinto virtuale a forma di T. Mentre ogni topo si muoveva lungo il lungo corridoio del labirinto, brevi “torri” visive apparivano sulle pareti di sinistra e destra. L’obiettivo dell’animale era semplice: alla fine, svoltare verso il lato che aveva mostrato più torri per ottenere una ricompensa. Risolvere questo compito richiede di sommare le torri per diversi secondi e mantenere quel totale in mente durante un ritardo finale prima di voltare. Mentre i topi addestrati eseguivano centinaia di queste prove, il team ha usato tecniche avanzate di imaging del calcio per registrare l’attività di oltre 14.000 neuroni in quattro regioni cerebrali coinvolte nella presa di decisione e nella memoria.
Da rampa liscia a onde correnti
Le teorie classiche, ispirate a registrazioni in scimmie e altri animali, propongono che singoli neuroni aumentino o diminuiscano gradualmente i loro tassi di scarica mentre l’evidenza si accumula verso una scelta. Ma in questi dati dei topi, quella storia non ci stava. Invece di rampa lente e persistenti, i neuroni nella corteccia, nell’ippocampo e nello striato tendevano a scaricare brevemente a turno, formando sequenze ordinate legate al progresso dell’animale lungo il labirinto e alla scelta finale. Una cellula poteva sparare principalmente a metà del corridoio quando il topo poi avrebbe svoltato a sinistra, mentre un’altra cellula sparava più tardi per le svolte a destra. Queste «sequenze selettive per la scelta» attraversavano ciascuna regione mentre la prova si svolgeva, mettendo in discussione l’idea che un singolo gruppo stabile di neuroni mantenga la variabile decisionale.
Due progetti di circuito per totali progressivi
Per spiegare come una sequenza mutevole di cellule potesse comunque portare un totale progressivo preciso dell’evidenza, gli autori hanno costruito e analizzato due famiglie di modelli di circuito neurale. Nel primo, chiamato «catene in competizione», l’evidenza è immagazzinata nella forza relativa dell’attività in due linee parallele di neuroni, una a favore della sinistra e una a favore della destra. Un segnale di gating dipendente dalla posizione assicura che solo una piccola coppia di neuroni—uno per ogni catena—sia attiva in ciascun punto del labirinto. Le torri in entrata spingono la coppia attiva verso un lato, e connessioni specializzate trasmettono quell’asimmetria accumulata alla coppia successiva man mano che l’animale si muove. In questo progetto, la quantità di evidenza è codificata in modo monotono: più evidenza a destra significa semplicemente attività più forte nella catena destra e più debole in quella sinistra. Nel secondo modello, un «bump gated dalla posizione», ogni posizione lungo il labirinto ospita un foglio di neuroni che collettivamente formano un compatto bump di attività lungo un asse dell’evidenza. Le torri spostano il bump a sinistra o a destra, come scorrere un indicatore lungo una scala. Man mano che il topo avanza, connessioni feedforward passano questo bump al foglio successivo, preservandone la posizione. Qui, i neuroni esatti attivi in un dato momento indicano il livello di evidenza, generando una sintonizzazione a picco stretto anziché ampie rampe.
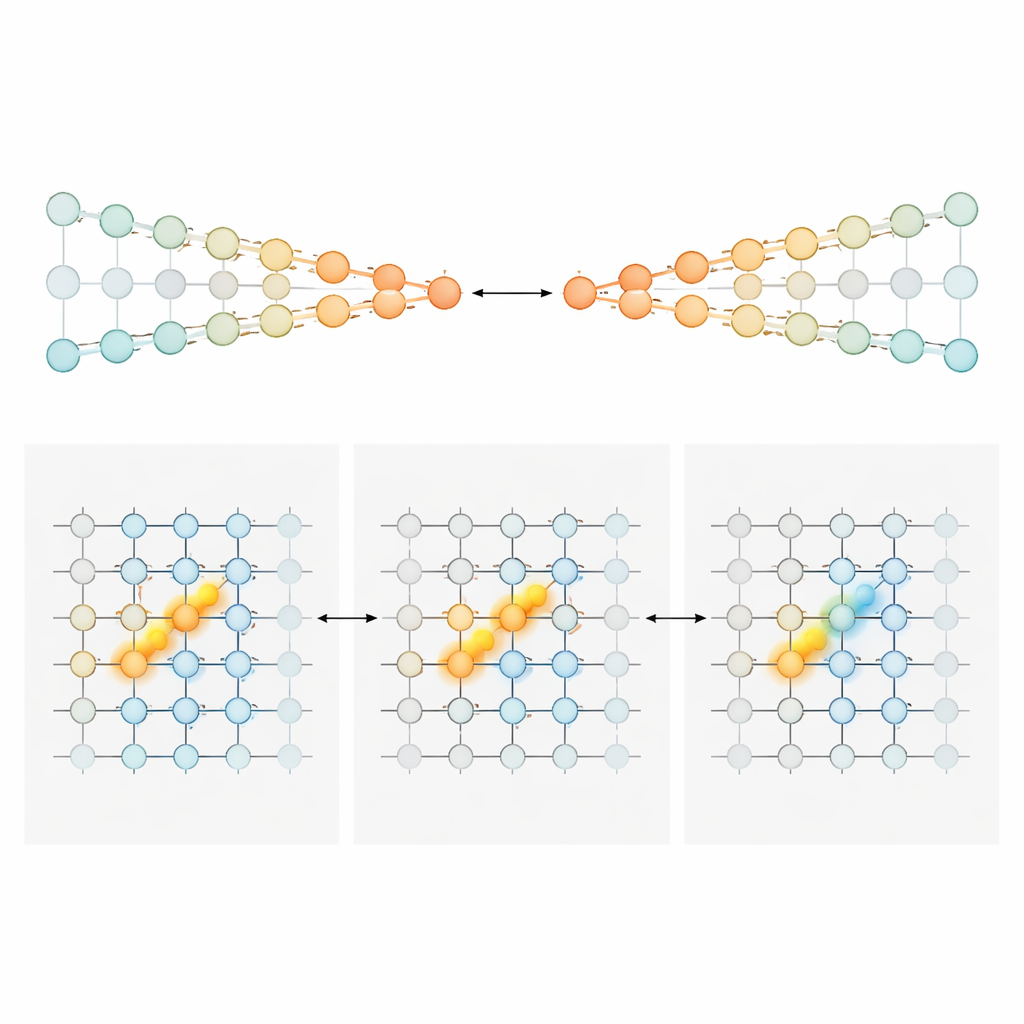
Zone cerebrali diverse, stili diversi di conteggio
Armati di questi progetti, il team è tornato alle registrazioni per chiedersi quali regioni assomigliassero di più a quale modello. Nella corteccia cingolata anteriore e nella corteccia retrospleniale—aree frontali e parietali implicate nella pianificazione e nella navigazione—la maggior parte dei neuroni mostrava una sensibilità ampia e per lo più unidirezionale all’evidenza. La loro attività tendeva ad aumentare costantemente con un supporto più forte per una scelta preferita, corrispondendo allo stile di codifica delle catene in competizione. All’inizio del periodo degli indizi, queste popolazioni trasportavano un segnale graduato su «quanto» più evidenza favorisse un lato. Man mano che il topo si avvicinava al punto di scelta, la risposta della popolazione si affilava, collassando gradualmente in un segnale più categorico, simile alla scelta. Per contrasto netto, i neuroni ippocampali tipicamente rispondevano in modo stretto e a forma di campana a valori specifici di evidenza, e cellule diverse ricoprivano l’intera gamma di evidenza. Questo schema è esattamente ciò che predice il modello del bump e si adatta al ruolo più ampio dell’ippocampo nel mappare dove si trova un animale e cosa è accaduto lungo il suo percorso. I neuroni dello striato, registrati principalmente durante il ritardo, tendevano a sparare in modo più tutto-o-nulla legato alla svolta finale, coerente con un ruolo a valle nella selezione dell’azione.
Cosa significa questo per comprendere le decisioni
Il messaggio centrale dello studio è che il cervello può usare più soluzioni di circuito per tenere traccia delle evidenze che si svolgono, anche all’interno di un singolo compito. Alcune regioni sembrano rappresentare l’evidenza come una graduale contesa tra pool di neuroni in competizione, adatta a confrontare opzioni e guidare una scelta finale. Altre, come l’ippocampo, rappresentano l’evidenza in modo più simile a una mappa, dove l’identità delle cellule attive segnala direttamente il conto corrente. Entrambi gli schemi si basano su sequenze di attività transitorie piuttosto che su scariche statiche, con connessioni strutturate che passano l’informazione lungo mentre l’«orologio» interno del compito avanza. Insieme, questi modelli e misurazioni suggeriscono che le nostre decisioni emergono da meccanismi flessibili e specifici per regione che possono sia sommare indizi rumorosi sia instradare il totale progressivo al posto giusto al momento giusto.
Citazione: Brown, L.S., Cho, J.R., Bolkan, S.S. et al. Neural circuit models for evidence accumulation through choice-selective sequences. Nat Commun 17, 4055 (2026). https://doi.org/10.1038/s41467-026-70267-9
Parole chiave: presa di decisione, accumulazione di prove, circuiti neurali, ippocampo, sequenze corticali