Clear Sky Science · es
Modelos de circuitos neuronales para la acumulación de evidencia mediante secuencias selectivas de elección
Cómo el cerebro suma silenciosamente las pistas
Las decisiones cotidianas, desde cruzar una calle hasta elegir un aperitivo, dependen de la capacidad del cerebro para sumar silenciosamente fragmentos de evidencia a lo largo del tiempo. Durante décadas, los neurocientíficos pensaron que esto lo realizaban grupos de neuronas que se mantenían activas de forma sostenida, como un indicador mental que se eleva lentamente hasta llegar a una decisión. Este estudio plantea qué ocurre cuando esa imagen clásica es incorrecta—cuando las células cerebrales disparan en patrones rápidos y cambiantes—y revela nuevos diseños de circuitos que el cerebro podría usar para llevar la cuenta de la evidencia conforme se despliega.
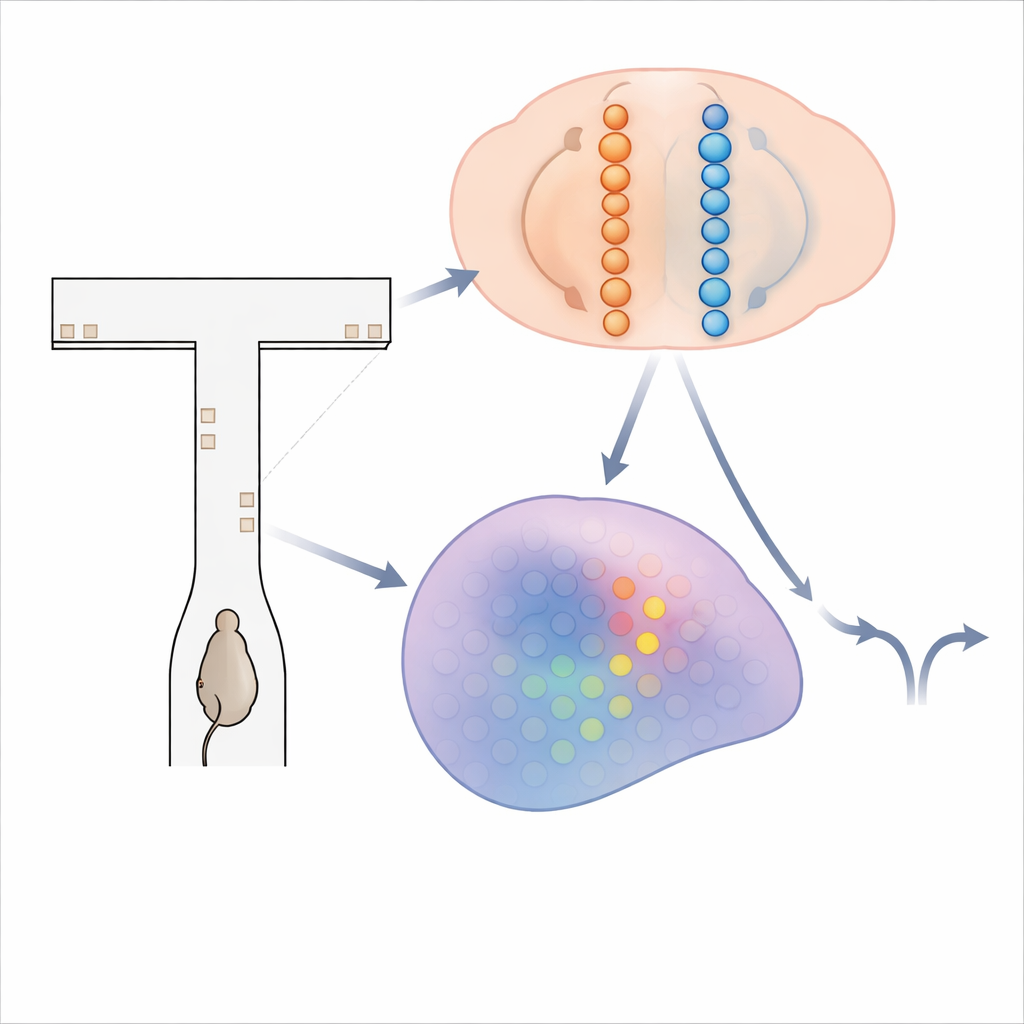
Un laberinto de pistas parpadeantes
Para investigar cómo el cerebro acumula evidencia, los investigadores usaron ratones que recorrían un laberinto virtual con forma de T. Mientras cada ratón avanzaba por el largo pasillo central, breves "torres" visuales aparecían en las paredes izquierda y derecha. La meta del animal era sencilla: al final, girar hacia el lado que había mostrado más torres para obtener una recompensa. Resolver esta tarea requiere sumar las torres durante varios segundos y mantener ese total en mente durante un retraso final antes de girar. Mientras los ratones entrenados realizaban cientos de estas pruebas, el equipo empleó técnicas avanzadas de registro por calcio para grabar la actividad de más de 14.000 neuronas en cuatro regiones cerebrales implicadas en la toma de decisiones y la memoria.
De rampas suaves a olas que compiten
Las teorías clásicas, inspiradas en registros en monos y otros animales, proponen que neuronas individuales aumentan o disminuyen gradualmente sus tasas de disparo a medida que la evidencia se acumula hacia una elección. Pero en estos datos de ratón, esa historia no encajó. En lugar de rampas lentas y persistentes, las neuronas en corteza, hipocampo y estriado tendían a disparar brevemente por turno, formando secuencias ordenadas vinculadas al avance del animal por el laberinto y a su elección final. Una célula podría activarse principalmente a mitad del pasillo cuando el ratón luego giraría a la izquierda, mientras que otra se activaba más tarde para giros a la derecha. Estas “secuencias selectivas por elección” barrían cada región conforme se desarrollaba la prueba, desafiando la idea de que un único grupo estable de neuronas sostiene la variable de decisión.
Dos planos de circuito para totales en marcha
Para explicar cómo una secuencia cambiante de células aún podría transportar un total acumulado preciso de evidencia, los autores construyeron y analizaron dos familias de modelos de circuitos neuronales. En el primero, llamado "cadenas en competencia", la evidencia se almacena en la fuerza relativa de la actividad en dos líneas paralelas de neuronas, una que favorece la izquierda y otra la derecha. Una señal de compuerta dependiente de la posición asegura que solo un pequeño par de neuronas—una de cada cadena—esté activo en cada punto del laberinto. Las torres entrantes empujan al par activo hacia un lado, y conexiones especializadas transmiten ese desequilibrio acumulado al siguiente par conforme el animal avanza. En este diseño, la cantidad de evidencia se codifica de forma monótona: más evidencia hacia la derecha simplemente significa mayor actividad en la cadena derecha y menor en la izquierda. En el segundo modelo, una "protuberancia condicionada por la posición", cada posición a lo largo del laberinto aloja una lámina de neuronas que colectivamente forman una protuberancia compacta de actividad a lo largo de un eje de evidencia. Las torres desplazan la protuberancia a la izquierda o a la derecha, como deslizar un marcador a lo largo de una escala. A medida que el ratón progresa, conexiones feedforward transfieren esta protuberancia a la siguiente lámina, preservando su ubicación. Aquí, las neuronas exactas activas en un momento dado indican el nivel de evidencia, produciendo una sintonía estrecha en forma de pico en lugar de rampas amplias.
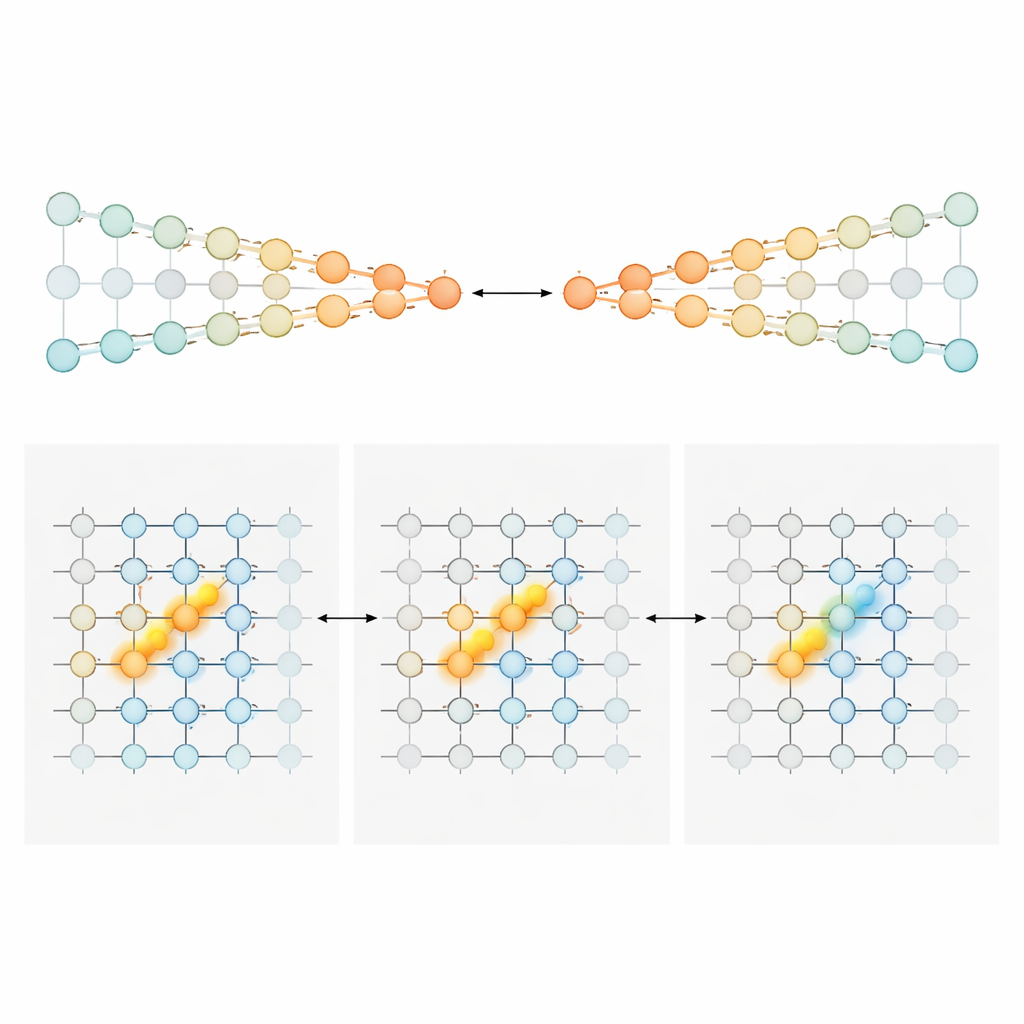
Diferentes áreas cerebrales, distintos estilos de contar
Armados con estos planos, el equipo volvió a los registros para preguntar qué regiones se parecían más a cada modelo. En la corteza cingulada anterior y en la corteza retrosplenial—áreas frontales y parecidas al lóbulo parietal implicadas en la planificación y la navegación—la mayoría de las neuronas mostraron una sensibilidad amplia y mayoritariamente unidireccional a la evidencia. Su actividad tendía a aumentar de forma sostenida con un apoyo más fuerte hacia la elección preferida, coincidiendo con el estilo de codificación de cadenas en competencia. Al inicio del periodo de señales, estas poblaciones portaban una señal graduada sobre "cuánto" más evidencia favorecía un lado. A medida que el ratón se acercaba al punto de elección, la respuesta poblacional se aguzaba, colapsando gradualmente hacia una señal más categórica, similar a una elección. En marcado contraste, las neuronas hipocampales respondían típicamente de forma estrecha y en forma de campana a valores específicos de evidencia, y diferentes células cubrían todo el rango de evidencia. Este patrón es exactamente lo que predice el modelo de protuberancia y encaja con el papel más amplio del hipocampo en mapear dónde está un animal y qué ha ocurrido a lo largo de su recorrido. Las neuronas del estriado, registradas principalmente durante el retraso, tendieron a disparar de manera más de todo-o-nada vinculada al giro eventual, coherente con un papel aguas abajo en la selección de la acción.
Qué significa esto para entender las decisiones
El mensaje central del estudio es que el cerebro puede usar múltiples trucos de circuito para llevar la cuenta de la evidencia que se despliega, incluso dentro de una única tarea. Algunas regiones parecen representar la evidencia como una lucha continua entre grupos de neuronas en competencia, bien adaptada para comparar opciones y dirigir una elección final. Otras, como el hipocampo, representan la evidencia de manera más tipo mapa, donde la identidad de las células activas señala directamente el conteo actual. Ambos esquemas se basan en secuencias de actividad transitoria en lugar de disparos estáticos, con conexiones cuidadosamente estructuradas que transmiten la información a medida que avanza el “reloj” interno de la tarea. En conjunto, estos modelos y mediciones sugieren que nuestras decisiones emergen de mecanismos flexibles y específicos por región que pueden tanto sumar pistas ruidosas como encaminar el total acumulado al lugar correcto en el momento adecuado.
Cita: Brown, L.S., Cho, J.R., Bolkan, S.S. et al. Neural circuit models for evidence accumulation through choice-selective sequences. Nat Commun 17, 4055 (2026). https://doi.org/10.1038/s41467-026-70267-9
Palabras clave: toma de decisiones, acumulación de evidencia, circuitos neuronales, hipocampo, secuencias corticales