Clear Sky Science · de
Neuronale Schaltkreis-Modelle zur Akkumulation von Evidenz durch wahlselektive Sequenzen
Wie das Gehirn leise Hinweise zusammenzählt
Alltägliche Entscheidungen, vom Überqueren einer Straße bis zur Wahl eines Snacks, beruhen auf der Fähigkeit des Gehirns, über die Zeit Stücke von Evidenz stillschweigend zu addieren. Jahrzehntelang gingen Neurowissenschaftler davon aus, dass dies von Gruppen von Neuronen geleistet wird, die dauerhaft aktiv bleiben – wie ein innerer Zähler, der langsam in Richtung einer Entscheidung ansteigt. Diese Studie fragt, was passiert, wenn dieses klassische Bild falsch ist – wenn Hirnzellen stattdessen in schnellen, wechselnden Mustern feuern – und zeigt neue Schaltungsdesigns auf, die das Gehirn möglicherweise nutzt, um die sich entwickelnde Evidenz zu verfolgen.
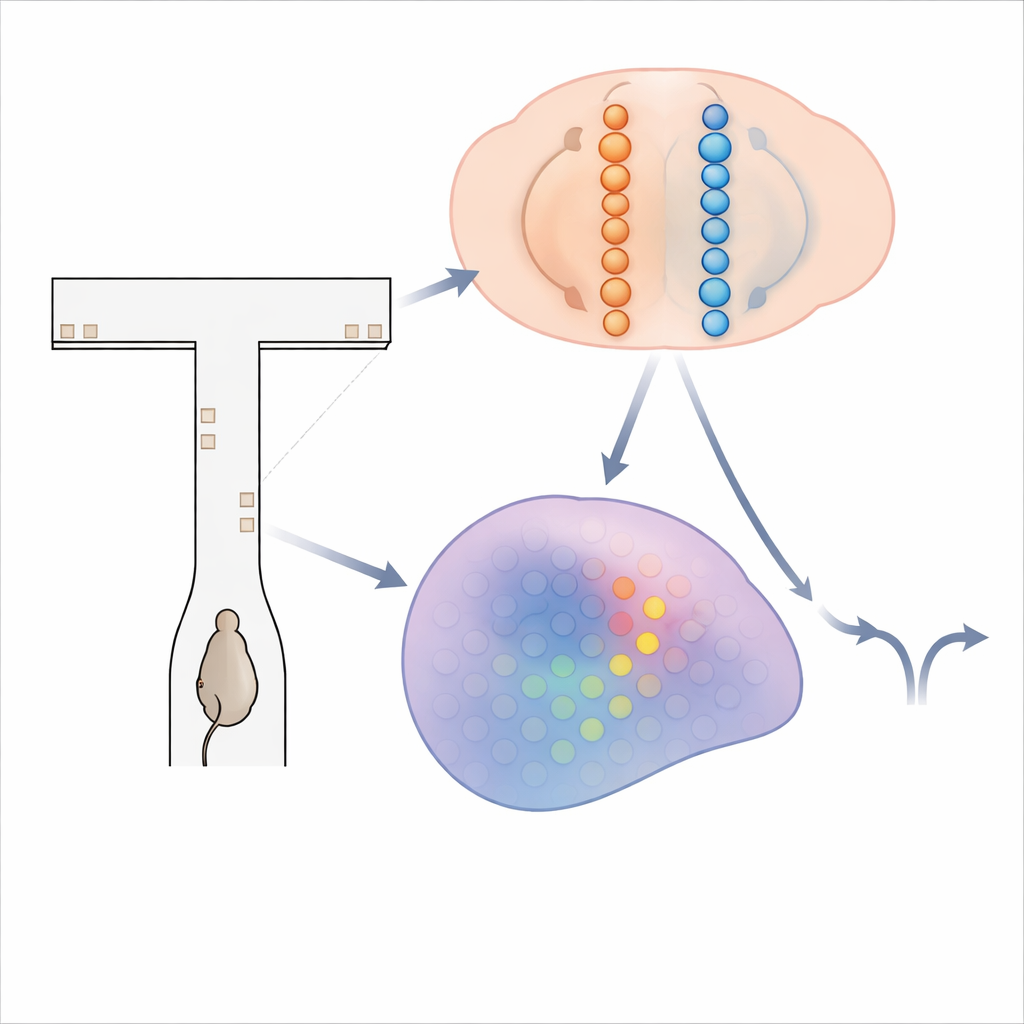
Ein Labyrinth flackernder Hinweise
Um zu untersuchen, wie das Gehirn Evidenz akkumuliert, setzten die Forscher Mäuse in ein virtuelles T-förmiges Labyrinth. Während jede Maus den langen Schenkel des Labyrinths entlanglief, erschienen kurz visuelle „Türme“ an den linken und rechten Wänden. Das Ziel des Tiers war einfach: Am Ende musste es sich zu der Seite wenden, die mehr Türme gezeigt hatte, um eine Belohnung zu erhalten. Die Lösung dieser Aufgabe erfordert das Aufaddieren der Türme über mehrere Sekunden und das Vorhalten dieser laufenden Summe während einer abschließenden Verzögerung vor der Wendung. Während die trainierten Mäuse Hunderte solcher Durchgänge absolvierten, nutzte das Team hochauflösende Calcium-Bildgebung, um die Aktivität von mehr als 14.000 Neuronen in vier an Entscheidungsfindung und Gedächtnis beteiligten Hirnregionen aufzuzeichnen.
Von glatten Rampen zu rasenden Wellen
Klassische Theorien, angeregt durch Aufzeichnungen bei Affen und anderen Tieren, schlagen vor, dass einzelne Neuronen ihre Feuerraten allmählich hoch- oder herunterfahren, während die Evidenz zugunsten einer Wahl akkumuliert wird. In diesen Mausdaten passte diese Darstellung jedoch nicht. Anstelle langsamer, persistenter Rampen feuerten Neuronen in Cortex, Hippocampus und Striatum eher kurz nacheinander und bildeten geordnete Sequenzen, die mit dem Fortgang der Maus durch das Labyrinth und mit ihrer späteren Wahl verknüpft waren. Eine Zelle konnte vorwiegend in der Mitte des Korridors feuern, wenn die Maus später nach links abbog, während eine andere Zelle später bei Rechtswendungen aktiv war. Diese „wahlselektiven Sequenzen“ zogen sich durch jede Region, während der Versuch ablief, und stellten die Vorstellung in Frage, eine einzelne stabile Gruppe von Neuronen halte die Entscheidungsvariable.
Zwei Schaltkreis-Entwürfe für laufende Summen
Um zu erklären, wie eine sich verschiebende Sequenz von Zellen dennoch eine präzise laufende Summe der Evidenz tragen kann, entwickelten und analysierten die Autor*innen zwei Familien neuronaler Schaltkreis-Modelle. Im ersten, den „konkurrierenden Ketten“, wird Evidenz in der relativen Stärke der Aktivität zweier paralleler Neuronenlinien gespeichert, eine für links, eine für rechts. Ein positionsabhängiges Gate-Signal sorgt dafür, dass zu jedem Punkt im Labyrinth nur ein kleines Paar von Neuronen – je eines aus jeder Kette – aktiv ist. Eintreffende Türme schieben das aktive Paar zugunsten einer Seite, und spezialisierte Verbindungen geben dieses akkumulierte Ungleichgewicht beim Voranschreiten an das nächste Paar weiter. In diesem Entwurf wird die Evidenzmenge monoton kodiert: Mehr rechtseitige Evidenz bedeutet einfach stärkere Aktivität in der rechten Kette und schwächere in der linken. Im zweiten Modell, einem „positionsgegateten Bump“, beherbergt jede Position entlang des Labyrinths ein Geflecht von Neuronen, das kollektiv eine kompakte Aktivitätskuppe entlang einer Evidenzachse bildet. Türme verschieben diese Kuppe nach links oder rechts, wie ein Marker, der auf einer Skala verschoben wird. Wenn die Maus voranschreitet, übergeben Feedforward-Verbindungen diese Kuppe an das nächste Geflecht und bewahren so ihre Lage. Hier geben die genau aktiven Neuronen zu einem Zeitpunkt den Evidenzstand an und liefern schmale, spitzenförmige Tunings statt breiter Rampen.
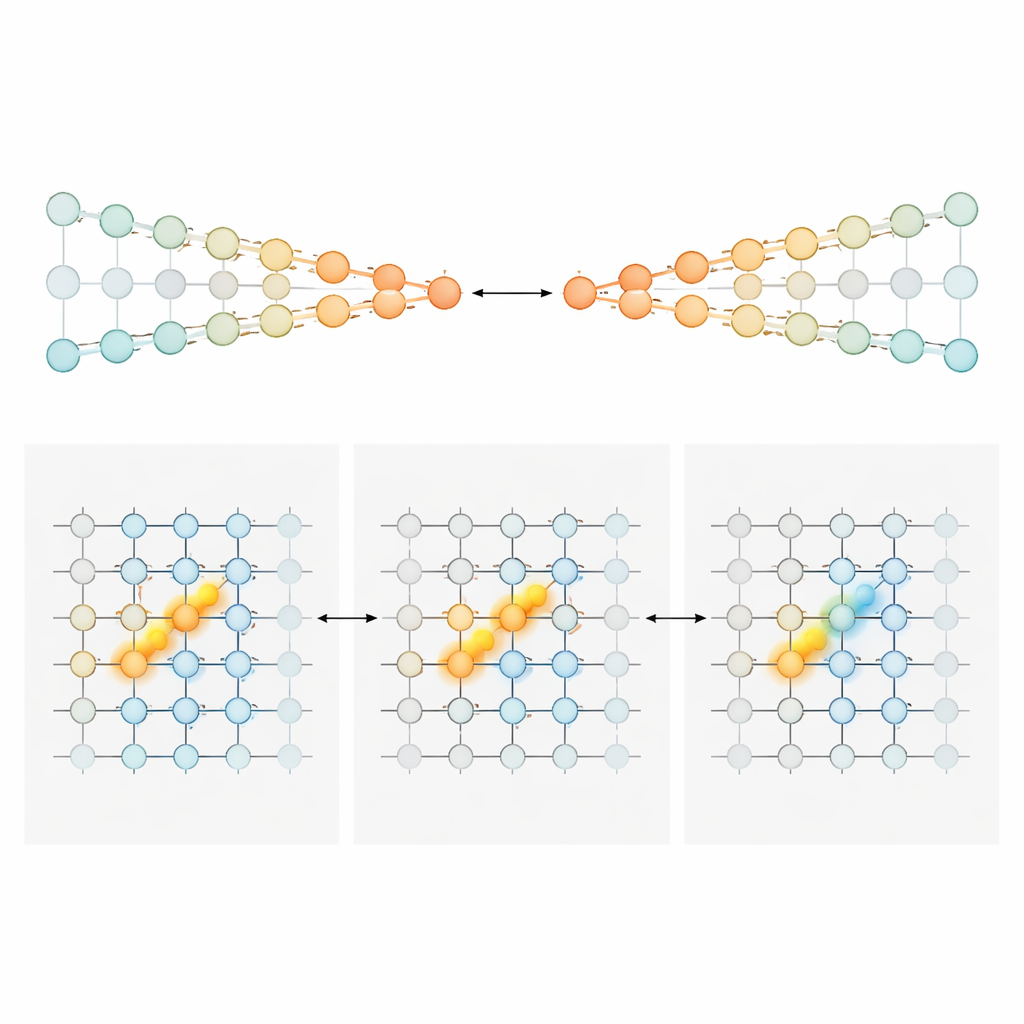
Verschiedene Hirnareale, verschiedene Zählstile
Mit diesen Entwürfen im Gepäck betrachtete das Team die Aufzeichnungen erneut, um zu fragen, welche Regionen eher welchem Modell entsprachen. Im anterioren cingulären Cortex und im retrosplenialen Cortex – frontale und parietalähnliche Bereiche, die an Planung und Navigation beteiligt sind – zeigten die meisten Neuronen eine breite, größtenteils eindirektionale Sensitivität gegenüber Evidenz. Deren Aktivität stieg tendenziell gleichmäßiger an, je stärker die Unterstützung für eine bevorzugte Wahl war, und entsprach damit dem Kodierungsstil der konkurrierenden Ketten. Früh in der Hinweisperiode trugen diese Populationen ein abgestuftes Signal darüber, „wie viel“ mehr Evidenz eine Seite begünstigte. Während die Maus sich dem Wendepunkt näherte, schärfte sich die Populationsantwort und brach allmählich zu einem kategorischeren, wahlähnlichen Signal zusammen. Ganz anders verhielten sich hippocampale Neuronen: Sie reagierten typischerweise eng und glockenförmig auf spezifische Evidenzwerte, wobei verschiedene Zellen das gesamte Evidenzspektrum abdeckten. Dieses Muster entspricht genau der Vorhersage des Bump-Modells und passt zu der weiteren Rolle des Hippocampus bei der Kartierung, wo sich ein Tier befindet und was entlang seines Weges geschehen ist. Striatale Neuronen, hauptsächlich während der Verzögerungsphase aufgezeichnet, feuerten eher alles-oder-nichts-ähnlich in Verbindung mit der späteren Wendung, was mit einer nachgelagerten Rolle bei der Aktionsauswahl vereinbar ist.
Was das für das Verständnis von Entscheidungen bedeutet
Die zentrale Botschaft der Studie ist, dass das Gehirn mehrere Schaltungstricks nutzen kann, um sich entwickelnde Evidenz zu verfolgen, selbst innerhalb einer einzigen Aufgabe. Manche Regionen scheinen Evidenz als einen glatten Tauziehen zwischen konkurrierenden Neuronengruppen darzustellen, gut geeignet zum Vergleichen von Optionen und zur Steuerung einer finalen Wahl. Andere, wie der Hippocampus, repräsentieren Evidenz eher kartenartig, wobei die Identität der aktiven Zellen direkt den aktuellen Stand anzeigt. Beide Schemata beruhen auf Sequenzen transitorischer Aktivität statt auf statischem Feuern, mit sorgfältig strukturierten Verbindungen, die Informationen weiterreichen, während die interne „Uhr“ der Aufgabe voranschreitet. Zusammen deuten diese Modelle und Messungen darauf hin, dass unsere Entscheidungen aus flexiblen, regionsspezifischen Mechanismen entstehen, die sowohl verrauschte Hinweise aufsummieren als auch die laufende Summe zur richtigen Zeit an den richtigen Ort leiten können.
Zitation: Brown, L.S., Cho, J.R., Bolkan, S.S. et al. Neural circuit models for evidence accumulation through choice-selective sequences. Nat Commun 17, 4055 (2026). https://doi.org/10.1038/s41467-026-70267-9
Schlüsselwörter: Entscheidungsfindung, Evidenzakkumulation, neurale Schaltkreise, Hippocampus, kortikale Sequenzen