Clear Sky Science · fr
Modèles de circuits neuronaux pour l’accumulation d’informations via des séquences sélectives de choix
Comment le cerveau additionne discrètement les indices
Les choix quotidiens, de traverser une rue à choisir une collation, reposent sur la capacité du cerveau à additionner silencieusement des bribes de preuves au fil du temps. Pendant des décennies, les neuroscientifiques ont pensé que cela se faisait par des groupes de neurones qui restaient activement soutenus, comme un compteur mental qui monte lentement vers une décision. Cette étude examine ce qui se passe lorsque ce tableau classique est faux — lorsque les cellules cérébrales tirent plutôt dans des motifs rapides et changeants — et révèle de nouveaux dessins de circuits que le cerveau pourrait utiliser pour suivre les preuves au fur et à mesure qu’elles se présentent.
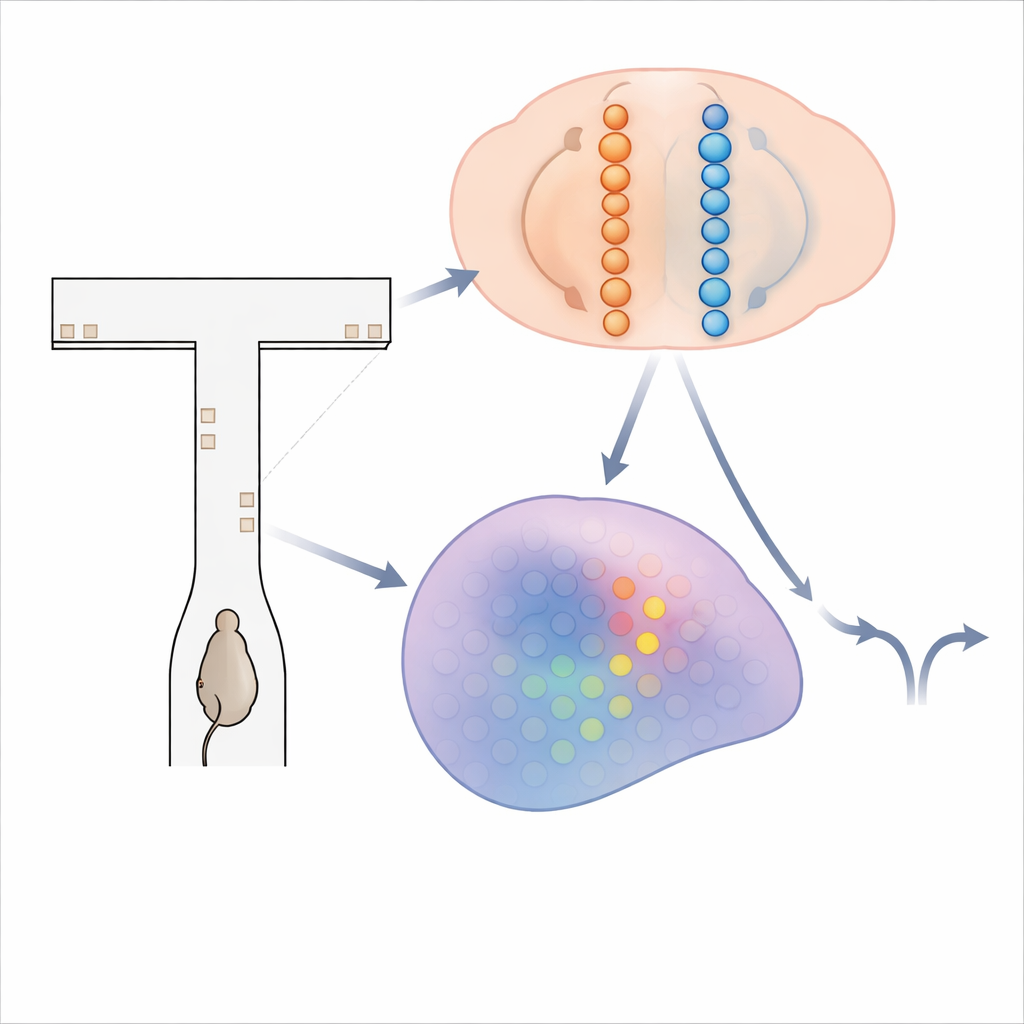
Un labyrinthe d’indices fugaces
Pour sonder la manière dont le cerveau accumule des preuves, les chercheurs ont utilisé des souris courant dans un labyrinthe virtuel en T. À mesure que chaque souris descendait la longue tige du labyrinthe, de brèves « tours » visuelles apparurent sur les murs gauche et droit. L’objectif de l’animal était simple : au bout, se tourner vers le côté qui avait montré le plus de tours pour gagner une récompense. Résoudre cette tâche exige d’additionner les tours sur plusieurs secondes et de conserver ce total en mémoire pendant un délai final avant de se diriger. Pendant que des souris entraînées effectuaient des centaines de ces essais, l’équipe a utilisé l’imagerie calcique avancée pour enregistrer l’activité de plus de 14 000 neurones à travers quatre régions cérébrales impliquées dans la prise de décision et la mémoire.
Des rampes lisses aux vagues en course
Les théories classiques, inspirées par des enregistrements chez le singe et d’autres animaux, proposent que des neurones isolés augmentent ou diminuent graduellement leur taux de décharge à mesure que les preuves s’accumulent en faveur d’un choix. Mais dans ces données de souris, ce récit ne coïncidait pas. Au lieu de rampes lentes et persistantes, les neurones du cortex, de l’hippocampe et du striatum avaient tendance à tirer brièvement à tour de rôle, formant des séquences ordonnées liées aux progrès de l’animal dans le labyrinthe et à son choix final. Une cellule pouvait tirer principalement à mi-chemin du couloir quand la souris se tournerait plus tard à gauche, tandis qu’une autre cellule tirait plus tard pour les virages à droite. Ces « séquences sélectives de choix » balayaient chaque région au fil de l’essai, remettant en cause l’idée qu’un groupe stable de neurones unique porte la variable de décision.
Deux plans de circuit pour des totaux courants
Pour expliquer comment une séquence changeante de cellules pourrait tout de même porter un total courant précis des preuves, les auteurs ont construit et analysé deux familles de modèles de circuits neuronaux. Dans le premier, appelé « chaînes concurrentes », les preuves sont stockées dans la force relative d’activité de deux lignes parallèles de neurones, l’une favorisant la gauche et l’autre la droite. Un signal de gating dépendant de la position veille à ce qu’une petite paire de neurones — une dans chaque chaîne — soit active à chaque point le long du labyrinthe. Les tours entrants poussent la paire active vers un côté, et des connexions spécialisées transmettent ce déséquilibre accumulé à la paire suivante au fur et à mesure que l’animal avance. Dans ce dispositif, la quantité de preuve est encodée de façon monotone : plus de preuves en faveur de la droite signifie simplement une activité plus forte dans la chaîne droite et plus faible dans la gauche. Dans le second modèle, un « bump position-gated » (bosse pilotée par la position), chaque position du labyrinthe héberge une couche de neurones qui forment collectivement une bosse compacte d’activité le long d’un axe de preuve. Les tours déplacent la bosse vers la gauche ou la droite, comme faire glisser un marqueur le long d’une échelle. À mesure que la souris avance, des connexions feedforward transmettent cette bosse à la couche suivante, préservant sa localisation. Ici, les neurones exacts actifs à un instant donné indiquent le niveau de preuve, produisant un réglage étroit en forme de pic plutôt que de larges rampes.
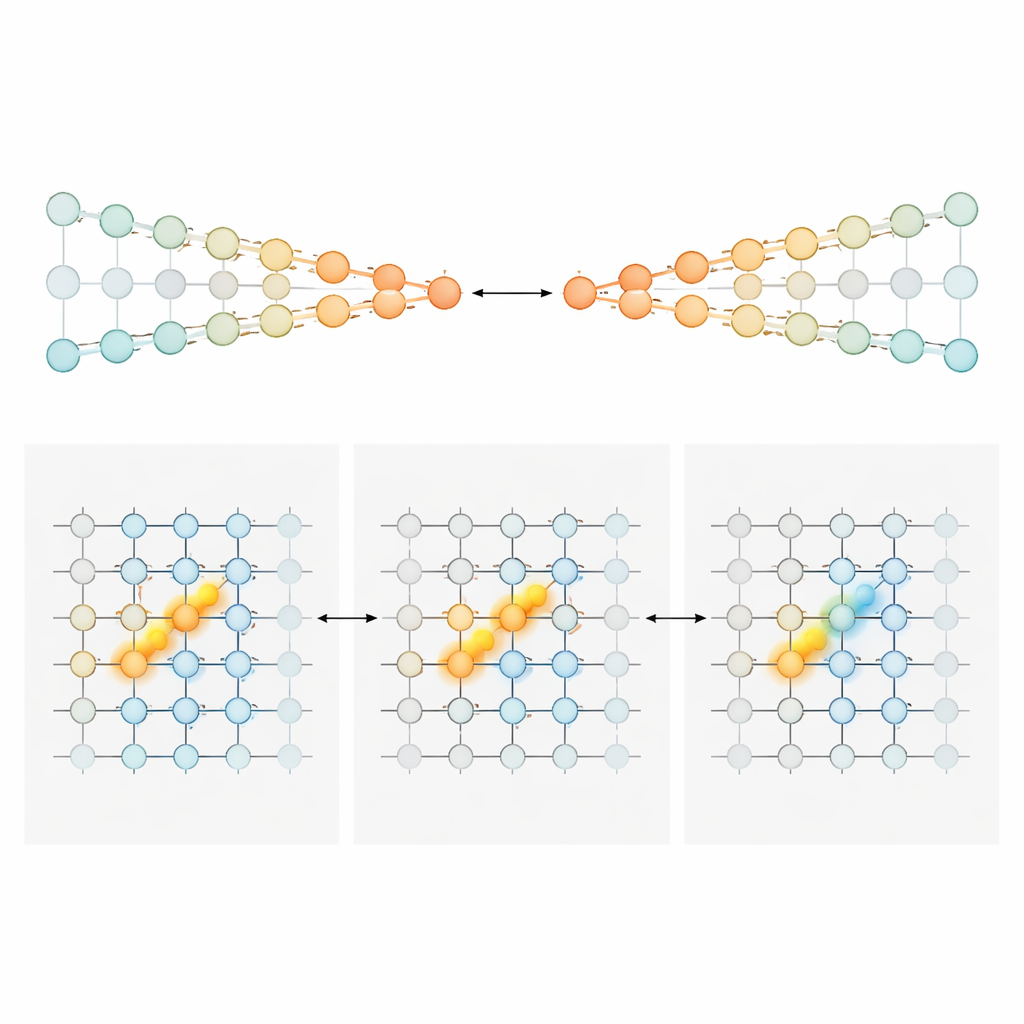
Différentes aires cérébrales, différents styles de comptage
Armée de ces plans, l’équipe est retournée aux enregistrements pour demander quelles régions ressemblaient le plus à quel modèle. Dans le cortex cingulaire antérieur et le cortex rétrosplénial — zones frontales et pariétales impliquées dans la planification et la navigation — la plupart des neurones montraient une sensibilité large, majoritairement unidirectionnelle, aux preuves. Leur activité avait tendance à augmenter de manière régulière avec un soutien plus fort en faveur d’un choix préféré, correspondant au style de codage des chaînes concurrentes. Au début de la période d’indices, ces populations portaient un signal gradué sur « combien » davantage de preuves favorisaient un côté. À l’approche du point de choix, la réponse de la population s’affinait, se condensant progressivement en un signal plus catégorique lié au choix. En contraste marqué, les neurones hippocampiques répondaient typiquement de façon étroite et en cloche à des valeurs d’évidence spécifiques, et différentes cellules couvraient l’ensemble de l’échelle des preuves. Ce schéma correspond exactement à ce que prédit le modèle de la bosse et s’accorde avec le rôle plus large de l’hippocampe dans la cartographie de la position d’un animal et de ce qui s’est produit le long de son trajet. Les neurones du striatum, enregistrés principalement pendant le délai, avaient tendance à tirer de manière plus tout-ou-rien liée au virage final, cohérente avec un rôle en aval dans la sélection d’action.
Ce que cela signifie pour la compréhension des décisions
Le message central de l’étude est que le cerveau peut utiliser plusieurs astuces de circuit pour suivre des preuves en cours, même au sein d’une même tâche. Certaines régions semblent représenter les preuves comme une lutte continue entre pools concurrents de neurones, bien adaptée pour comparer des options et provoquer un choix final. D’autres, comme l’hippocampe, représentent les preuves de manière plus cartographique, où l’identité des cellules actives signale directement le total courant. Les deux schémas reposent sur des séquences d’activités transitoires plutôt que sur des décharges statiques, avec des connexions soigneusement structurées qui transmettent l’information au fur et à mesure que « l’horloge » interne de la tâche avance. Ensemble, ces modèles et mesures suggèrent que nos décisions émergent de mécanismes flexibles, spécifiques aux régions, capables à la fois d’additionner des indices bruités et d’acheminer le total courant au bon endroit au bon moment.
Citation: Brown, L.S., Cho, J.R., Bolkan, S.S. et al. Neural circuit models for evidence accumulation through choice-selective sequences. Nat Commun 17, 4055 (2026). https://doi.org/10.1038/s41467-026-70267-9
Mots-clés: prise de décision, accumulation de preuves, circuits neuronaux, hippocampe, séquences corticales