Clear Sky Science · en
Neural circuit models for evidence accumulation through choice-selective sequences
How the brain quietly tallies up clues
Everyday choices, from crossing a street to picking a snack, depend on the brain’s ability to silently add up bits of evidence over time. For decades, neuroscientists thought this was done by groups of neurons that stayed steadily active, like a mental meter slowly rising toward a decision. This study asks what happens when that classic picture is wrong—when brain cells instead fire in rapid, shifting patterns—and reveals new circuit designs the brain may use to keep track of evidence as it unfolds.
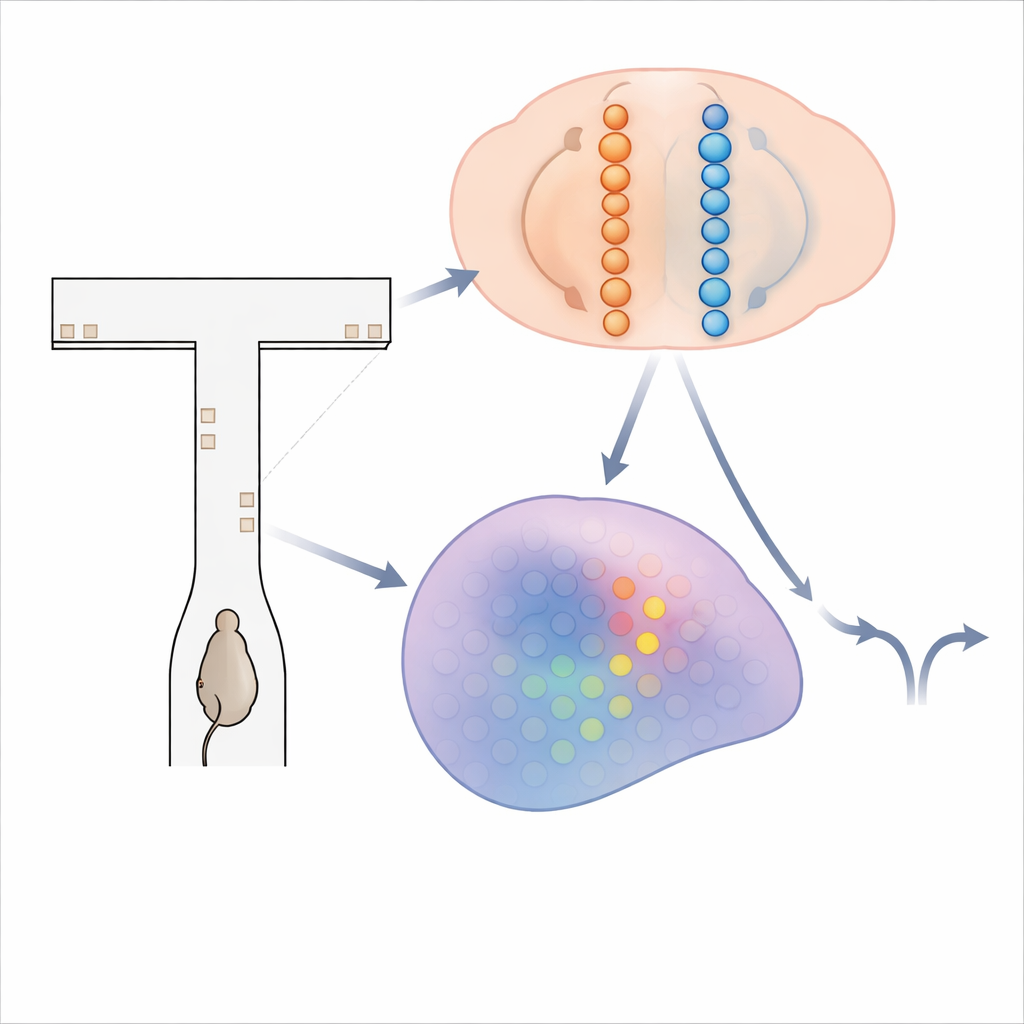
A maze of flickering clues
To probe how the brain accumulates evidence, the researchers used mice running through a virtual T-shaped maze. As each mouse moved down the long stem of the maze, brief visual “towers” popped up on the left and right walls. The animal’s goal was simple: at the far end, turn toward the side that had shown more towers to earn a reward. Solving this task requires adding up towers over several seconds and holding that running total in mind during a final delay before turning. While trained mice performed hundreds of these trials, the team used advanced calcium imaging to record activity from more than 14,000 neurons across four brain regions involved in decision making and memory.
From smooth ramps to racing waves
Classic theories, inspired by recordings in monkeys and other animals, propose that single neurons gradually ramp their firing rates up or down as evidence accumulates toward one choice. But in these mouse data, that story did not fit. Instead of slow, persistent ramps, neurons in cortex, hippocampus, and striatum tended to fire briefly in turn, forming orderly sequences tied to the animal’s progress along the maze and to its eventual choice. A cell might fire mainly halfway down the corridor when the mouse would later turn left, while another cell fired later for right turns. These “choice-selective sequences” swept across each region as the trial unfolded, challenging the idea that a single stable group of neurons holds the decision variable.
Two circuit blueprints for running totals
To explain how a shifting sequence of cells could still carry a precise running total of evidence, the authors built and analyzed two families of neural circuit models. In the first, called “competing chains,” evidence is stored in the relative strength of activity in two parallel lines of neurons, one favoring left and one favoring right. A position-dependent gating signal ensures that only a small pair of neurons—one from each chain—are active at each point along the maze. Incoming towers nudge the active pair toward one side, and specialized connections pass that accumulated imbalance forward to the next pair as the animal moves. In this design, the amount of evidence is encoded monotonically: more rightward evidence simply means stronger activity in the right chain and weaker in the left. In the second model, a “position-gated bump,” each position along the maze hosts a sheet of neurons that collectively form a compact bump of activity along an evidence axis. Towers shift the bump left or right, like sliding a marker along a scale. As the mouse advances, feedforward connections hand off this bump to the next sheet, preserving its location. Here, the exact neurons active at a given time indicate the evidence level, yielding narrow, peak-shaped tuning rather than broad ramps.
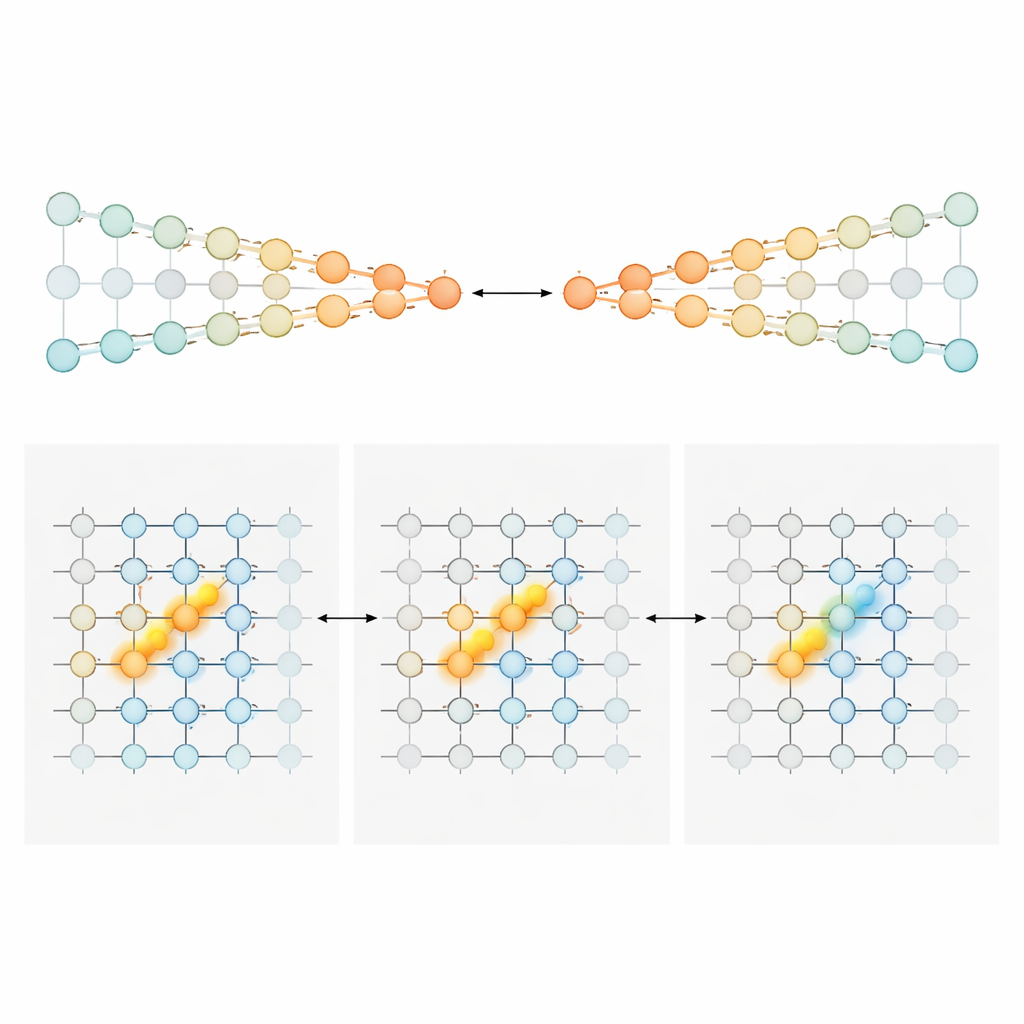
Different brain areas, different counting styles
Armed with these blueprints, the team turned back to the recordings to ask which regions looked more like which model. In anterior cingulate cortex and retrosplenial cortex—frontal and parietal-like areas involved in planning and navigation—most neurons showed broad, mostly one-directional sensitivity to evidence. Their activity tended to rise steadily with stronger support for a preferred choice, matching the competing-chains style of coding. Early in the cue period, these populations carried a graded signal about “how much” more evidence favored one side. As the mouse approached the choice point, the population response sharpened, gradually collapsing into a more categorical, choice-like signal. In stark contrast, hippocampal neurons typically responded in a narrow, bell-shaped fashion to specific evidence values, and different cells tiled the full evidence range. This pattern is exactly what the bump model predicts and fits with the hippocampus’s broader role in mapping where an animal is and what has happened along its path. Striatal neurons, recorded mainly during the delay, tended to fire in a more all-or-none way tied to the eventual turn, consistent with a downstream role in action selection.
What this means for understanding decisions
The study’s central message is that the brain may use multiple circuit tricks to keep track of unfolding evidence, even within a single task. Some regions appear to represent evidence as a smooth tug-of-war between competing pools of neurons, well suited for comparing options and driving a final choice. Others, like the hippocampus, represent evidence in a more map-like fashion, where the identity of the active cells directly signals the current tally. Both schemes rely on sequences of transient activity rather than static firing, with carefully structured connections passing information along as the internal “clock” of the task advances. Together, these models and measurements suggest that our decisions emerge from flexible, region-specific mechanisms that can both add up noisy clues and route the running total to the right place at the right time.
Citation: Brown, L.S., Cho, J.R., Bolkan, S.S. et al. Neural circuit models for evidence accumulation through choice-selective sequences. Nat Commun 17, 4055 (2026). https://doi.org/10.1038/s41467-026-70267-9
Keywords: decision making, evidence accumulation, neural circuits, hippocampus, cortical sequences