Clear Sky Science · pl
Modele obwodów nerwowych dla kumulacji dowodów poprzez sekwencje selektywne względem wyboru
Jak mózg cicho sumuje wskazówki
Codzienne decyzje, od przejścia przez ulicę po wybór przekąski, zależą od zdolności mózgu do dyskretnego dodawania kolejnych przesłanek w czasie. Przez dekady neurolodzy zakładali, że odbywa się to za pomocą grup neuronów utrzymujących stałą aktywność, niczym mentalny wskaźnik powoli rosnący w kierunku decyzji. To badanie pyta, co się dzieje, gdy ten klasyczny obraz jest fałszywy — gdy komórki mózgowe zamiast tego wyładowują się w szybkich, zmieniających się wzorcach — i ujawnia nowe projekty obwodów, które mózg może wykorzystywać, by śledzić dowody w miarę ich pojawiania się.
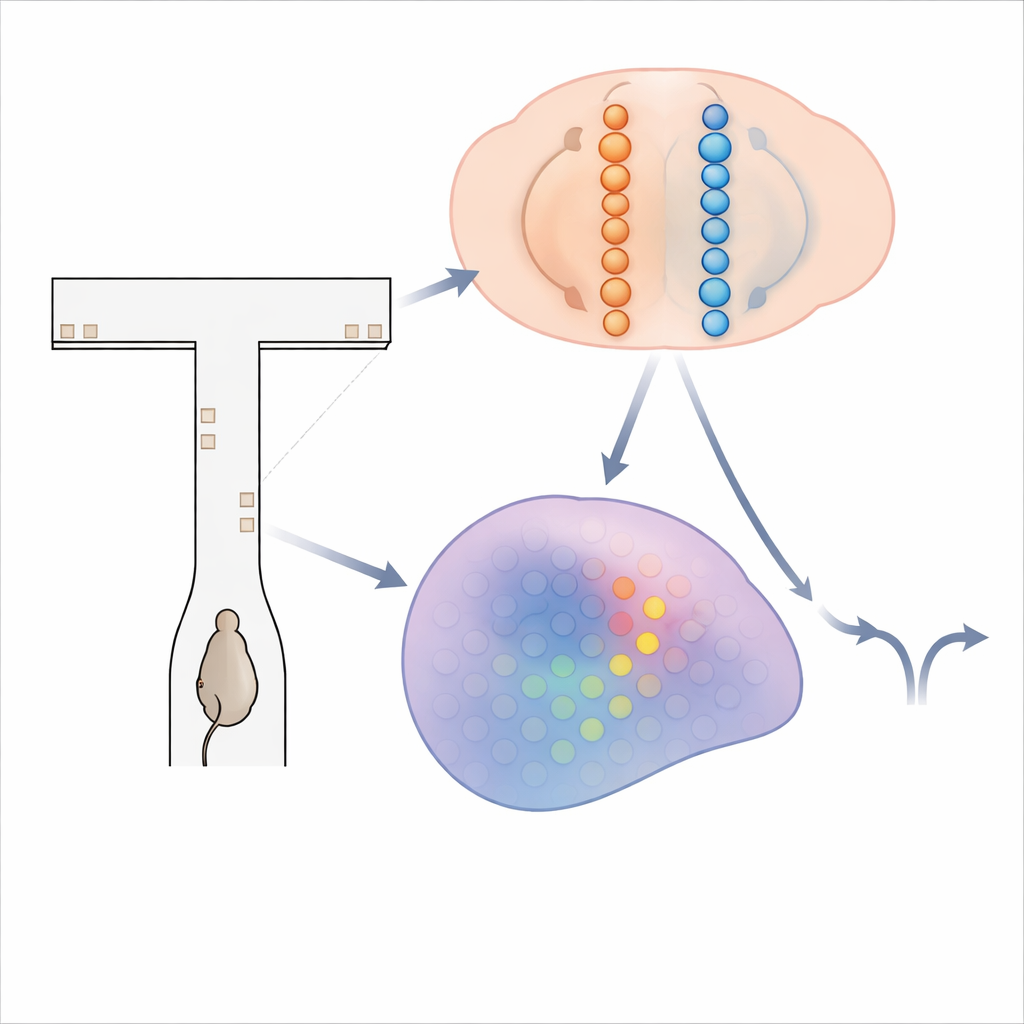
Labirynt migoczących wskazówek
Aby sprawdzić, jak mózg kumuluje dowody, badacze użyli myszy biegających w wirtualnym labiryncie w kształcie litery T. Gdy mysz poruszała się w dół długiego ramienia, krótkie wizualne „wieże” pojawiały się po lewej i prawej stronie ścian. Celem zwierzęcia było proste: na końcu skręcić w stronę, która pokazywała więcej wież, by otrzymać nagrodę. Rozwiązanie tego zadania wymaga zsumowania wież rozciągniętych na kilka sekund i utrzymania tej bieżącej sumy w pamięci podczas końcowego opóźnienia przed skrętem. Podczas gdy wyuczone myszy wykonywały setki takich prób, zespół użył zaawansowanego obrazowania wapniowego, aby zarejestrować aktywność ponad 14 000 neuronów w czterech obszarach mózgu zaangażowanych w podejmowanie decyzji i pamięć.
Od gładkich nachyleń do ścigających się fal
Klasyczne teorie, inspirowane zapisami u małp i innych zwierząt, zakładają, że pojedyncze neurony stopniowo zwiększają lub zmniejszają częstość wyładowań w miarę kumulowania dowodów na korzyść jednego wyboru. Ale w tych danych od myszy ten obraz nie pasował. Zamiast wolnych, trwałych nachyleń, neurony w korze, hipokampie i prążkowiu miały tendencję do krótkotrwałego wyładowywania się kolejno, tworząc uporządkowane sekwencje związane z postępem zwierzęcia w labiryncie i jego ostatecznym wyborem. Komórka mogła wyładowywać się głównie w połowie korytarza, gdy mysz później skręcała w lewo, podczas gdy inna komórka aktywowała się później przy skrętach w prawo. Te „sekwencje selektywne względem wyboru” przesuwały się przez każdy region w miarę przebiegu próby, kwestionując pomysł, że stała grupa neuronów utrzymuje zmienną decyzyjną.
Dwa projekty obwodów dla bieżących sum
Aby wyjaśnić, jak zmieniająca się sekwencja komórek mogła nadal nieść precyzyjną bieżącą sumę dowodów, autorzy zbudowali i przeanalizowali dwie rodziny modeli obwodów nerwowych. W pierwszym, zwanym „konkurencyjnymi łańcuchami”, dowody są przechowywane w względnej sile aktywności dwóch równoległych linii neuronów, jednej faworyzującej lewo i drugiej prawo. Sygnał bramkujący zależny od pozycji zapewnia, że w każdym punkcie labiryntu aktywna jest tylko mała para neuronów — po jednym z każdego łańcucha. Napływające wieże przesuwają aktywną parę w stronę jednej strony, a wyspecjalizowane połączenia przekazują to nagromadzone przesunięcie do następnej pary w miarę ruchu zwierzęcia. W tym rozwiązaniu ilość dowodów kodowana jest monotonicznie: silniejsza przewaga prawej strony oznacza po prostu silniejszą aktywność w prawym łańcuchu i słabszą w lewym. W drugim modelu, „pozycjonowanym garbie”, każda pozycja wzdłuż labiryntu mieści arkusz neuronów, które wspólnie tworzą zwartą górkę aktywności wzdłuż osi dowodów. Wieże przesuwają garb w lewo lub prawo, jak przesuwanie znacznika na skali. Gdy mysz posuwa się naprzód, połączenia feedforward przekazują ten garb do następnego arkusza, zachowując jego położenie. Tutaj dokładne neurony aktywne w danym momencie wskazują poziom dowodów, dając wąskie, szczytowe strojenie zamiast szerokich nachyleń.
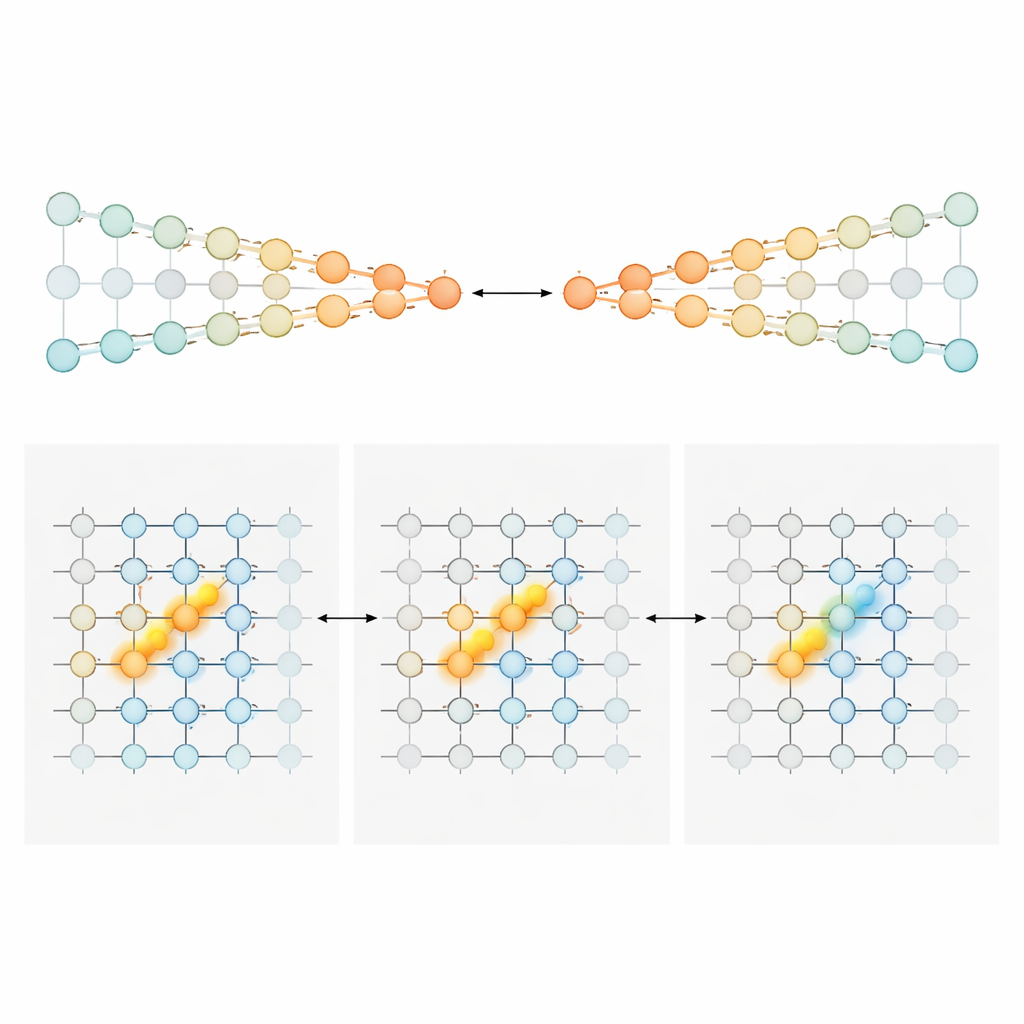
Różne obszary mózgu, różne style liczenia
Uzbrojeni w te projekty, autorzy wrócili do nagrań, by sprawdzić, które regiony bardziej przypominają który model. W korze obręczy przedniej i korze retrosplenialnej — obszarach przypominających przednią i ciemieniową część zaangażowanych w planowanie i nawigację — większość neuronów wykazywała szeroką, głównie jednostronną czułość na dowody. Ich aktywność miała tendencję do stopniowego wzrostu przy silniejszym wsparciu dla preferowanego wyboru, co odpowiada stylowi kodowania konkurencyjnych łańcuchów. Wczesne w okresie wskazówek populacje te niosły stopniowany sygnał o „ile” więcej dowodów sprzyja jednej stronie. Gdy mysz zbliżała się do punktu wyboru, odpowiedź populacji się zawężała, stopniowo zapadając w bardziej kategoryczny sygnał związany z wyborem. W rażącym kontraście neurony hipokampa zazwyczaj odpowiadały w wąski, dzwonowaty sposób na konkretne wartości dowodów, a różne komórki pokrywały cały zakres dowodów. Ten wzorzec jest dokładnie tym, co przewiduje model garbu i pasuje do szerszej roli hipokampa w mapowaniu, gdzie znajduje się zwierzę i co wydarzyło się wzdłuż jego trasy. Neurony prążkowia, rejestrowane głównie podczas opóźnienia, miały tendencję do wyładowań bardziej typu wszystko-albo-nic związanych z ostatecznym skrętem, co jest zgodne z rolą następczą w wyborze działania.
Co to znaczy dla rozumienia decyzji
Główne przesłanie badania jest takie, że mózg może używać wielu trików obwodowych, by śledzić rozwijające się dowody, nawet w ramach jednego zadania. Niektóre regiony wydają się reprezentować dowody jako płynne przeciąganie się sił między konkurującymi pulami neuronów, dobrze przystosowane do porównywania opcji i pchnięcia ostatecznego wyboru. Inne, jak hipokamp, reprezentują dowody w bardziej mapopodobny sposób, gdzie tożsamość aktywnych komórek bezpośrednio sygnalizuje bieżącą sumę. Oba schematy opierają się na sekwencjach przejściowej aktywności, a nie na statycznym wyładowaniu, z precyzyjnie ukształtowanymi połączeniami przekazującymi informacje w miarę przesuwania się wewnętrznego „zegar” zadania. Razem te modele i pomiary sugerują, że nasze decyzje wyłaniają się z elastycznych, specyficznych dla regionów mechanizmów, które potrafią zarówno dodawać zaszumione wskazówki, jak i kierować bieżącą sumę we właściwe miejsce we właściwym czasie.
Cytowanie: Brown, L.S., Cho, J.R., Bolkan, S.S. et al. Neural circuit models for evidence accumulation through choice-selective sequences. Nat Commun 17, 4055 (2026). https://doi.org/10.1038/s41467-026-70267-9
Słowa kluczowe: podejmowanie decyzji, kumulacja dowodów, obwody nerwowe, hipokamp, sekwencje korowe