Clear Sky Science · zh
使用协同进化模型在细胞中识别与工程化高性能花蕈病毒蛋白酶
为科学与医学打造更锐利的分子剪刀
现代生物学常依赖于微小的分子“剪刀”——蛋白酶——在恰当的位置切割蛋白质。这些切割可以开启或关闭细胞过程,帮助纯化实验室制备的蛋白,或驱动工程化的基因电路。然而,对于研究中广泛使用的一个大型病毒蛋白酶家族,其切割能力与精确性尚未被全面绘制。本研究提出了一种以数据为驱动的方法,用于发现、预测并重新设计这些蛋白酶,使其在人体细胞中更高效且更具选择性,从而为更清洁的生物技术应用乃至靶向破坏有害细胞打开了可能性。
为什么病毒蛋白切割酶很重要
这项工作聚焦于NIa蛋白酶——来自感染植物的花蕈病毒的酶。该家族的一员,烟草蚀斑病毒蛋白酶(TEVp),因能识别特定的七氨基酸序列并在确定位置切割,成为蛋白工程中的常用工具。但TEVp只是超过3,800个相关蛋白酶中的一个,这些酶的能力大多尚未被探索。如果科学家能系统地理解哪个蛋白酶切割哪个序列,以及微小序列变化如何影响活性,他们就能为实验替换更优的“剪刀”、在细胞中构建更复杂的合成电路,并设计仅对疾病相关突变起反应的蛋白酶。
从自然模式中学习
为了解决这一问题,作者收集了3,817对天然的花蕈病毒蛋白酶及其对应底物,内容不仅包括核心的七位切割位点,还涵盖了周围的氨基酸。他们随后构建了ProSSpeC——一个计算模型,用来寻找蛋白酶与底物之间的协同进化模式,即在进化过程中共同变化以维持良好契合的位置。借助一个受物理启发的评分方案,模型为每一对蛋白酶–底物分配一个特异性分数:分数越有利(越负),该对越有可能高效切割。通过剔除源自总体相似性而非真实相互作用的模式,ProSSpeC聚焦于真正对识别和切割位点至关重要的特征。
在人体细胞中验证预测
团队接着检验这些分数是否能预测活细胞中的行为。他们在人体细胞中设计了一种荧光检测体系:成功切割会重组分裂的红色荧光蛋白,产生被绿色报告蛋白归一化的明亮信号。测试数十种蛋白酶–底物组合后发现,具有更强ProSSpeC分数的组合倾向于产生更亮的信号,且天然蛋白酶通常偏好其自身的天然目标序列。在15种蛋白酶与15种底物共225种组合中,计算分数与测得的荧光有良好相关性,并能准确区分会切割与不会切割的组合,即便仅考虑七氨基酸核心基序时亦然。
逐个构件微调切割
由于ProSSpeC以单个氨基酸为分辨率,作者利用它来探索当目标序列中仅改变一个构件会发生什么。对于多对蛋白酶–底物,他们预测出应能增强或削弱切割的突变,然后构建这些变体并测量其活性。模型分数的变化与荧光变化密切对应,证实该模型能预见单个残基如何改变性能。模型还强调了核心位点周围更宽序列背景的重要性:将简单重复的基序替换为天然的20氨基酸邻域常常能将切割效率提升数倍,这支持了由进化塑造的侧翼残基对识别精确调节的观点。
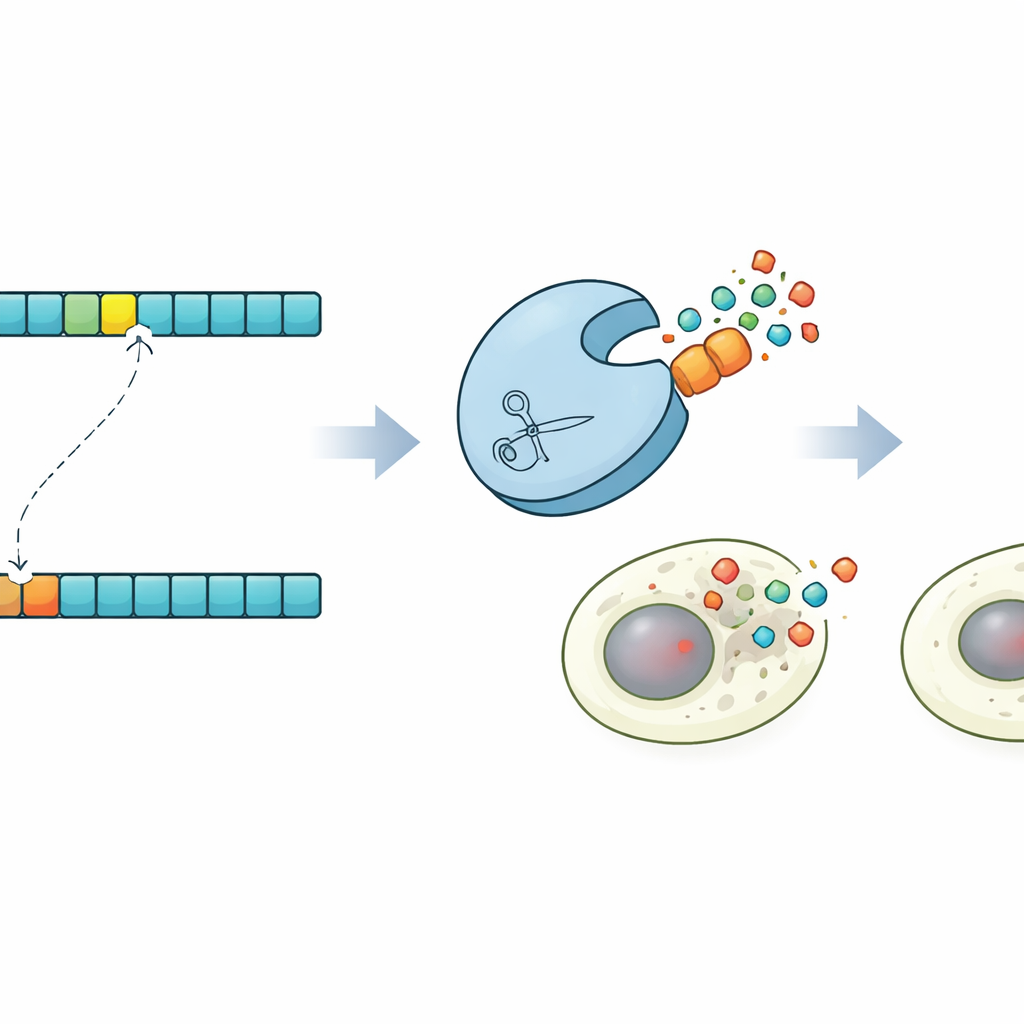
按指令触发程序性细胞死亡
为展示这种控制可以实现的功能,研究人员在人体细胞中设计了一个引人注目的演示。利用他们的预测,他们找到了一个能切割略微突变目标位点但不会切割原始位点的蛋白酶。随后他们构建了携带正常或突变位点的Caspase-3(程序性细胞死亡的关键执行者)变体。在混合细胞群体中,工程化的蛋白酶仅在携带突变序列的细胞中选择性激活Caspase-3,触发这些细胞的凋亡而不伤及邻近细胞。该“synpoptosis”电路表明,基于协同进化指导的蛋白酶设计可用于检测单个氨基酸差异并将其转换为细胞生死的决策。
对未来的意义
对非专业读者而言,关键的信息是作者已将散落在蛋白序列中的进化线索转化为一种实用的分子剪刀设计工具。ProSSpeC不仅能找到性能优于实验室常用工具如TEVp的蛋白酶,还能解释酶与底物之间哪些接触最重要,即便这些接触相距较远。尽管对于与天然序列相差甚远的序列存在局限,模型已经为研究者提供了一种浏览与重新设计数千种病毒蛋白酶的方法,以实现更干净的切割与定制的特异性。从长远看,此类工具可助力构建更智能的细胞治疗、更好的诊断手段,以及能够像CRISPR改变DNA那样以高精度编辑细胞内蛋白质网络的可编程系统。
引用: Lim Suan, M.B., Ziegler, C., Syed, Z. et al. Identification and engineering of highly functional potyviral proteases in cells using co-evolutionary models. Nat Commun 17, 3257 (2026). https://doi.org/10.1038/s41467-026-69961-5
关键词: 蛋白酶工程, 协同进化建模, 合成生物学, 程序性细胞死亡, 蛋白质特异性