Clear Sky Science · zh
ATPγS 大幅削弱驱动动蛋白步进的偏向机制
细胞货运工如何保持方向性
在每个细胞内部,名为动力蛋白的小型蛋白“行者”沿着微观轨道搬运货物,参与细胞的构建、修复和分裂。像高速公路上的卡车一样,这些行者必须在拉载重物时仍主要向前行驶。本研究提出了一个看似简单的问题:如果我们稍微改变它们的燃料,动力蛋白向前行走的能力会发生什么变化?答案揭示了一种意外的“停—走”状态,帮助马达保持正确航向。
一种分子行者及其常见燃料
Kinesin-1 是一种两足的马达蛋白,在交错于细胞的刚性纤维——微管上行走。每一步大约8纳米,由富含能量的分子 ATP 提供能量。动力蛋白的一个头牢牢抓住轨道,另一个头向前摆动,两个头以手递手的方式交替前进。在反向力作用下——比如负载向后拉——该马达仍然偏好向前迈步,这要归功于一种内部的“偏向”机制,使向前运动比向后滑动更可能发生。
用更慢的近亲替换 ATP
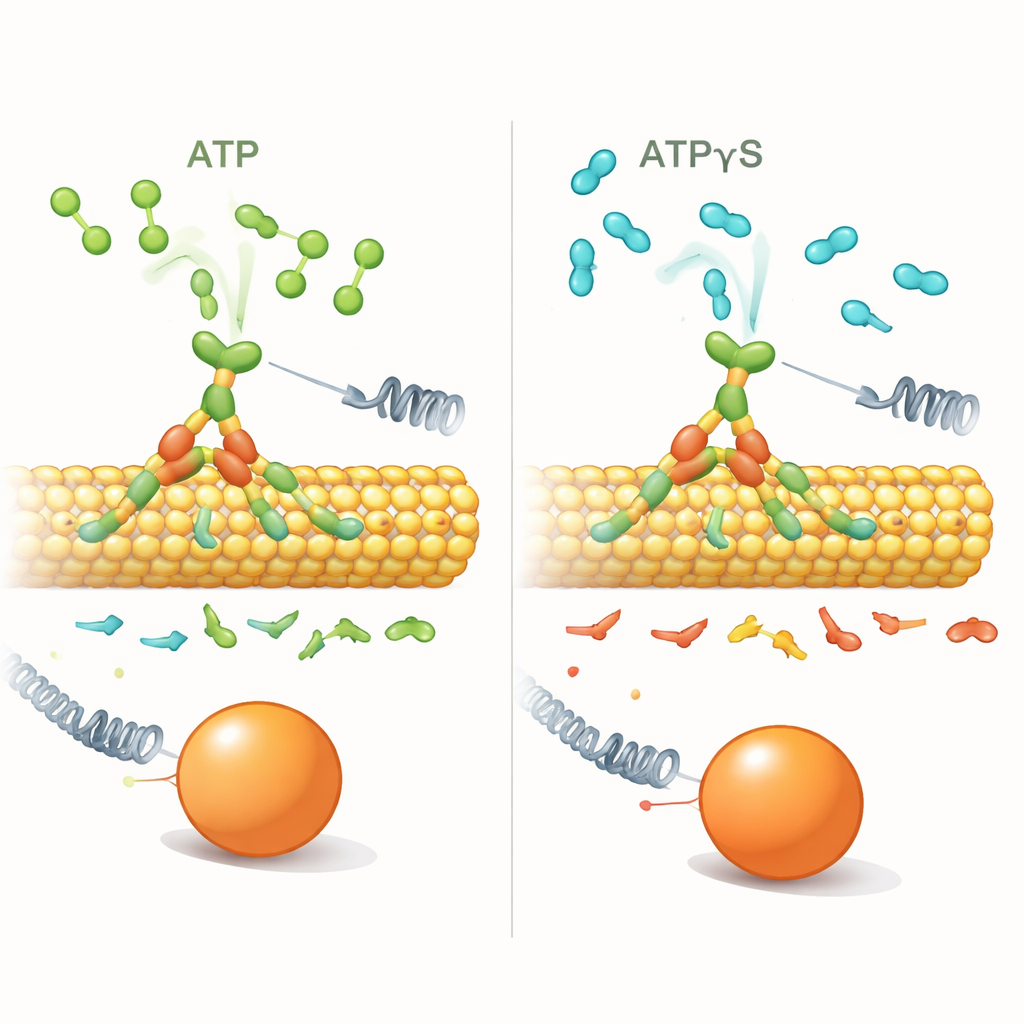
为了探究这一偏向是如何起作用的,研究者用 ATPγS 替换了 ATP,这是一种几乎相同但被马达分解得更慢的分子。借助单分子光学陷阱——可以抓住连接到单个动力蛋白的小珠的激光镊子——他们测量了在不同负载和燃料条件下,动力蛋白前进与后退步的发生频率。在低浓度 ATPγS(1 微摩尔)下,马达的行为与使用 ATP 时大体相同:其有过程性地向前行走,只有在负载增加时后退事件才变得更常见。
当过多缓慢燃料破坏偏向时
在高浓度 ATPγS(1 毫摩尔)下,情况发生了明显变化。动力蛋白仍在微管上行走并达到相似的停滞力,但此时即便在低负载下,也更频繁地出现短的 8 纳米后退步。前进与后退步的比率——在正常情况下随负载增加会急剧下降——变得几乎平坦:负载几乎不再起作用。与此同时,每一步前的等待时间——停留时间(dwell)——变得很长(约半秒或更长),并且对施加力的依赖很弱,这与 ATP 条件下停留时间随负载显著增长的情况不同。在 ATPγS 中,大多数后退是整洁的单步后退,而不是长距离“滑移”,这暗示着不同的运动机制。
一个隐藏的停顿态浮出水面
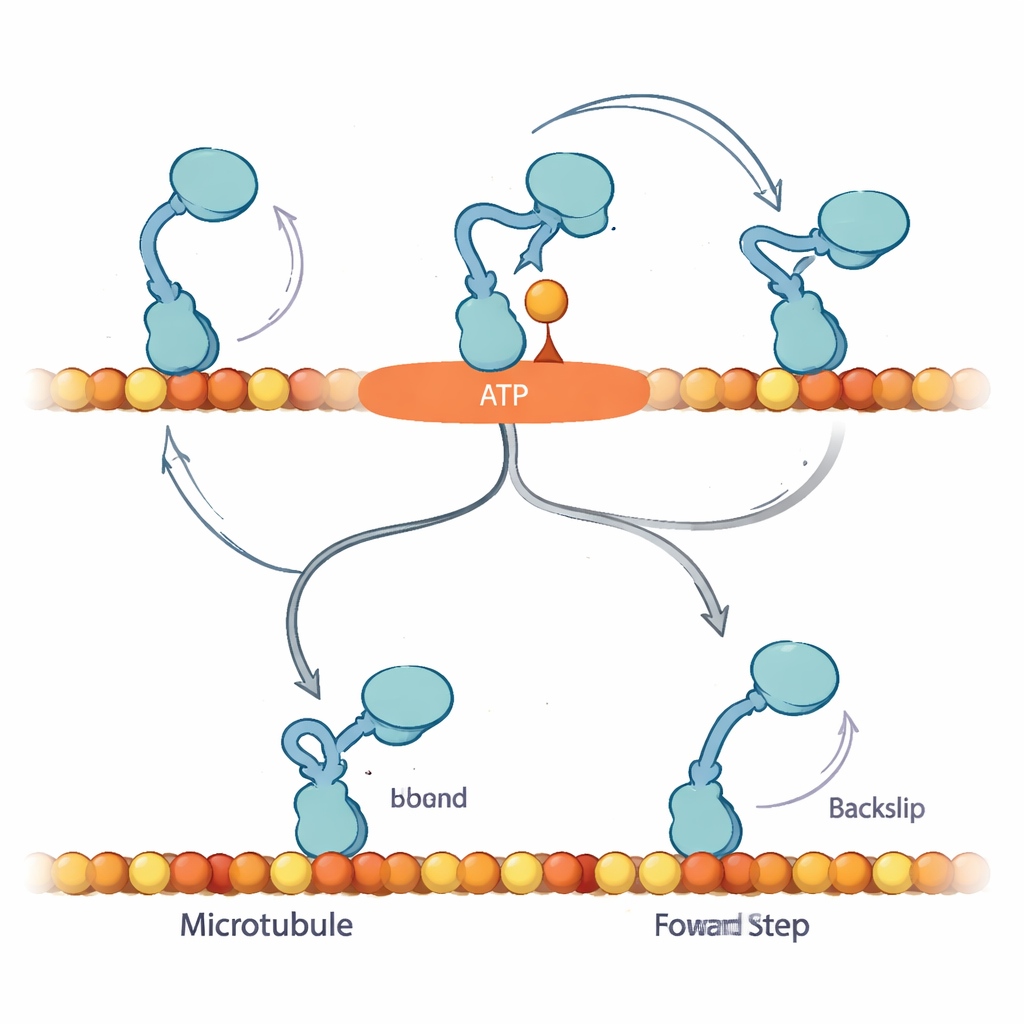
为了解释这些模式,作者提出,当核苷酸首次结合时,动力蛋白进入一个先前未被识别的“等待-构象异构化”(Await-Isomerisation,AI)态。在该状态下,ATP(或 ATPγS)已结合,马达已准备好迈步,但一段关键的结构片段——颈链(neck linker)——尚未锁定到位,且活性位点尚未准备好水解燃料。从 AI 态出发,自由头可以扩散,并且理论上可以结合到轨道的前方或后方。在正常 ATP 条件下,AI 态是短暂的:它很快转换为“闭合”态,颈链对接、随之发生水解,游离头被引导到前方位点,强烈偏向向前推进。大量 ATPγS 存在时,这一转换被延缓,AI 态被过度占据,从而打开了一条旁路,使马达更频繁地发生真实的 8 纳米后退步。
为什么可控的停顿很重要
这些发现表明,动力蛋白的偏向机制依赖于在不消耗燃料的情况下安全地停留在 AI 态,等待颈链对接以引导游离头到下一个前方结合位点。在有负载且使用 ATP 时,向前迈步与燃料水解紧密耦合:如果马达未能及时完成向前步伐,可能会向后滑动,但这相对少见。在 ATPγS 条件下,延长的 AI 态暴露出一个隐藏的后退通路,减少了向前的偏向,但仍允许总体向前移动。对于非专业读者来说,关键结论是:这个微小的马达并非只是消耗燃料然后盲目前进;它利用一个内建的“候车室”——可控的停顿——来决定是前进、停留还是后退。微妙地改变燃料揭示了这一隐藏逻辑,显示细胞如何通过微调运动来保持分子交通在应有的方向上流动,即使在受力情况下亦是如此。
引用: Karnawat, V., Toleikis, A., Carter, N.J. et al. ATPγS substantially defeats the biasing mechanism for kinesin steps. Nat Commun 17, 2891 (2026). https://doi.org/10.1038/s41467-026-69573-z
关键词: 动力蛋白(kinesin)马达, 分子马达, ATP 类似物, 单分子生物物理学, 微管运输