Clear Sky Science · pl
ATPγS w znacznym stopniu niweluje mechanizm faworyzujący kroki kinezyny
Jak komórkowi przewoźnicy ładunków zachowują kierunek
W każdej komórce drobne białkowe „chodzące” cząsteczki zwane kinezynami przewożą ładunki wzdłuż mikroskopijnych torów, pomagając w budowie, naprawie i podziale komórki. Jak ciężarówki na autostradzie, te „chodzące” muszą poruszać się w przeważającej mierze do przodu, nawet gdy ciągną ciężkie ładunki. To badanie stawia przewrotnie proste pytanie: co dzieje się ze zdolnością kinezyny do poruszania się do przodu, jeśli nieco zmienimy jej paliwo? Odpowiedź ujawnia niespodziewany stan pauzy i ruszania, który pomaga silnikowi utrzymać kurs.
Cząsteczkowy „wędrowiec” i jego zwykłe paliwo
Kinezyna-1 to dwuoczna cząsteczka motorowa, która „chodzi” po mikrotubulach — sztywnych włóknach przecinających komórkę. Każdy krok ma około 8 nanometrów i napędzany jest przez energetycznie bogate ATP. Jedna głowa kinezyny trzyma się mocno toru, podczas gdy druga wymachuje naprzód, a obie na przemian wykonują ruch typu „ręka za ręką”. Pod przeciwną siłą — jak przyciąganie obciążenia w tył — silnik wciąż faworyzuje kroki do przodu, dzięki wewnętrznemu mechanizmowi „uprzedzającemu”, który zwiększa prawdopodobieństwo ruchu naprzód względem cofnięć.
Wymiana ATP na wolniej działający odpowiednik
Aby zbadać, jak działa to faworyzowanie, badacze zastąpili ATP przez ATPγS, niemal identyczną cząsteczkę, którą silnik rozkłada znacznie wolniej. Przy użyciu pojedynczymolekularnych pułapek optycznych — pęset laserowych, które mogą trzymać kulkę przyczepioną do jednej kinezyny — mierzyli, jak często kinezyna wykonuje kroki do przodu względem kroków w tył w różnych obciążeniach i warunkach paliwowych. Przy niskich stężeniach ATPγS (1 mikromolarnym) zachowanie silnika było bardzo podobne do tego z ATP: poruszał się procesywnie do przodu, a zdarzenia cofnięć stawały się częstsze dopiero wraz ze wzrostem obciążenia.
Gdy zbyt dużo wolnego paliwa rozbija uprzedzenie
Obraz dramatycznie się zmienił przy wysokim stężeniu ATPγS (1 milimolarnym). Kinezyna wciąż wykonywała kroki po mikrotubuli i osiągała podobne siły hamujące, ale krótkie 8-nanometrowe kroki w tył stały się znacznie częstsze nawet przy niskich obciążeniach. Stosunek kroków naprzód do kroków w tył, który normalnie gwałtownie spada wraz ze wzrostem obciążenia, stał się niemal płaski: obciążenie miało niewielkie znaczenie. Równocześnie czas oczekiwania przed każdym krokiem — czas przestoju — był długi (około pół sekundy lub więcej) i słabo zależał od przyłożonej siły, w odróżnieniu od ATP, gdzie czasy przestoju silnie rosły wraz z obciążeniem. W ATPγS większość ruchów w tył to były schludne, jednopunktowe cofnięcia, a nie długie „poślizgi”, co sugeruje inny mechanizm ruchu.
Na światło dzienne wychodzi ukryty stan pauzy
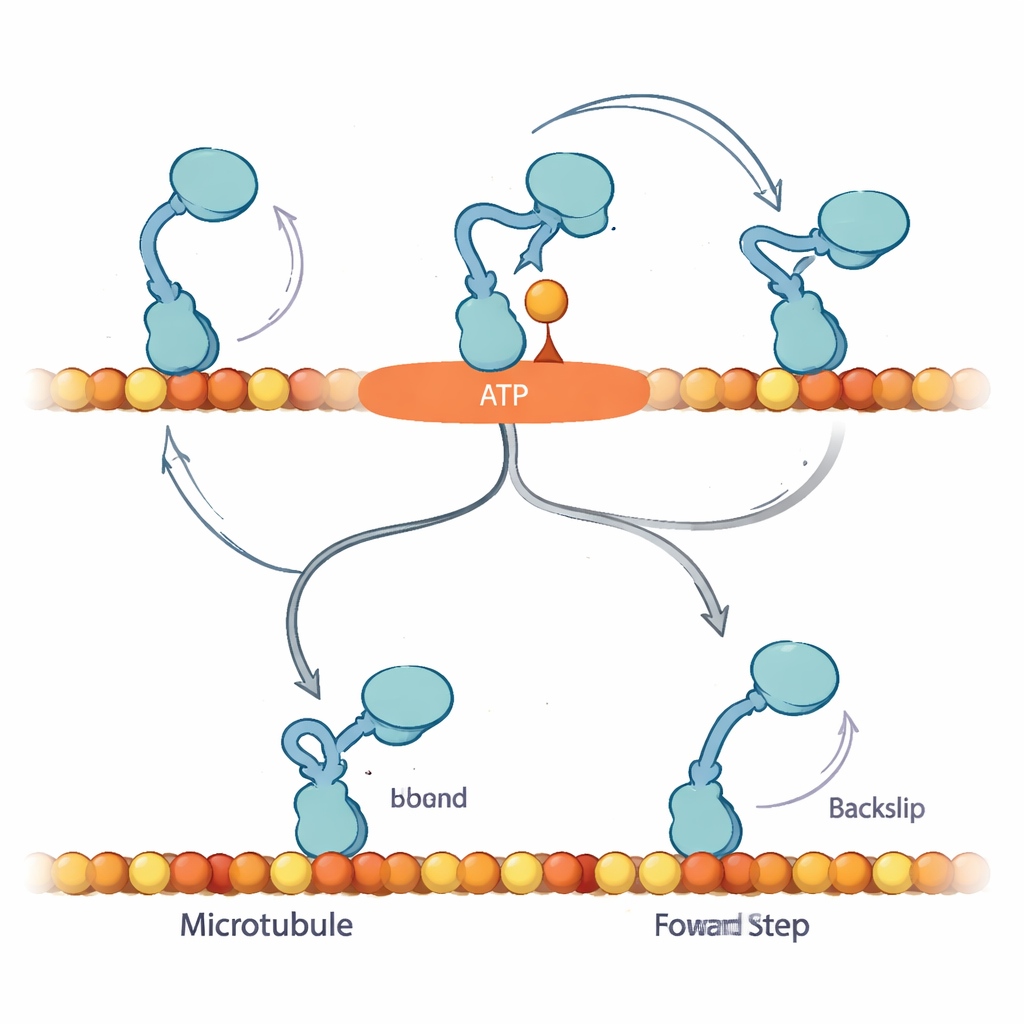
Aby wyjaśnić te wzorce, autorzy proponują, że po związaniu nukleotydu kinezyna wchodzi w wcześniej nieuznany stan „Await-Isomerisation” (AI). W tym stanie ATP (lub ATPγS) jest związane i motor jest przygotowany do kroku, ale kluczowy segment strukturalny, łącznik szyjny (neck linker), nie zablokował się jeszcze, a miejsce aktywne nie jest gotowe do rozkładu paliwa. Z AI wolna głowa dyfunduje i w zasadzie może związać się zarówno do przodu, jak i do tyłu na torze. W normalnym ATP stan AI jest krótkotrwały: szybko przechodzi w stan „zamknięty”, w którym łącznik szyjny dokuje, przebiega hydroliza, a wolna głowa jest skierowana do przedniego miejsca wiązania, co zdecydowanie faworyzuje ruch naprzód. Przy obfitym ATPγS ta konwersja jest spowolniona i AI jest nadmiernie obsadzony, otwierając boczną ścieżkę, w której silnik częściej wykonuje prawdziwe 8-nanometrowe kroki w tył.
Dlaczego kontrolowana pauza ma znaczenie
Wyniki sugerują, że mechanizm faworyzowania kinezyny opiera się na zdolności bezpiecznego zawieszenia się w stanie AI bez spalania paliwa, oczekując na dokowanie łącznika szyjnego, które skieruje wolną głowę do następnego przedniego miejsca wiązania. Pod obciążeniem z ATP kroki do przodu i rozkład paliwa są ściśle sprzężone: jeśli silnik nie ukończy kroku naprzód na czas, może się cofnąć, ale jest to stosunkowo rzadkie. W ATPγS przedłużony stan AI odsłania ukrytą drogę cofnięcia, zmniejszając preferencję dla ruchu naprzód, choć wciąż dopuszczając ruch netto do przodu. Dla laika kluczowe przesłanie jest takie, że ten maleńki silnik nie po prostu spala paliwo i idzie do przodu; wykorzystuje wbudowaną poczekalnię — kontrolowaną pauzę — by zdecydować, czy posunąć się naprzód, zostać, czy się cofnąć. Subtelna zmiana paliwa ujawnia tę ukrytą logikę, pokazując, jak komórki dopracowują ruch, by utrzymać przepływ molekularnego ruchu we właściwym kierunku, nawet pod obciążeniem.
Cytowanie: Karnawat, V., Toleikis, A., Carter, N.J. et al. ATPγS substantially defeats the biasing mechanism for kinesin steps. Nat Commun 17, 2891 (2026). https://doi.org/10.1038/s41467-026-69573-z
Słowa kluczowe: silnik kinezynowy, motory molekularne, analog ATP, biologia jednego cząsteczki, transport po mikrotubulach