Clear Sky Science · fr
ATPγS compromet sensiblement le mécanisme d'orientation des pas du kinésine
Comment les transporteurs cellulaires gardent le cap
À l’intérieur de chaque cellule, de minuscules « marcheurs » protéiques appelés kinésines déplacent des cargaisons le long de pistes microscopiques, participant à la construction, la réparation et la division de la cellule. À l’instar de camions sur une autoroute, ces marcheurs doivent avancer principalement vers l’avant, même lorsqu’ils tirent des charges lourdes. Cette étude pose une question apparemment simple : que devient la capacité du kinésine à avancer si l’on modifie légèrement son carburant ? La réponse révèle un état de pause et de reprise inattendu qui aide le moteur à rester sur la bonne trajectoire.
Un marcheur moléculaire et son carburant habituel
Le kinésine-1 est une protéine motrice bipède qui se déplace le long des microtubules, des filaments rigides qui parcourent la cellule. Chaque pas mesure environ 8 nanomètres et est alimenté par la molécule riche en énergie ATP. Une tête du kinésine reste fermement attachée à la piste pendant que l’autre se projette vers l’avant, les deux alternant dans un mouvement de type « main sur main ». Sous une force opposée — comme une charge tirant vers l’arrière — le moteur privilégie néanmoins les pas vers l’avant, grâce à un mécanisme interne de « biais » qui rend le mouvement vers l’avant plus probable que les glissements en arrière.
Remplacer l’ATP par un quasi-doubleur plus lent
Pour sonder le fonctionnement de ce biais, les chercheurs ont remplacé l’ATP par l’ATPγS, une molécule presque identique que le moteur hydrolyse beaucoup plus lentement. À l’aide de pièges optiques monocentriques — des pinces laser capables de retenir une bille attachée à un unique kinésine — ils ont mesuré la fréquence des pas avant versus arrière sous différentes charges et conditions de carburant. À faible concentration d’ATPγS (1 micromolaire), le moteur se comportait presque comme avec l’ATP : il avançait de manière processive, les événements de recul n’étant plus fréquents qu’à mesure que la charge augmentait.
Quand trop de carburant lent brise le biais
Le tableau a changé radicalement à haute concentration d’ATPγS (1 millimolaire). Le kinésine continuait à se déplacer le long du microtubule et atteignait des forces d’arrêt similaires, mais de courts pas arrière de 8 nanomètres devinrent beaucoup plus fréquents, même à faibles charges. Le ratio pas avant/pas arrière, qui décroît normalement fortement avec l’augmentation de la charge, devint presque plat : la charge importait peu. Parallèlement, le temps d’attente avant chaque pas — la période de dwell — était long (environ une demi-seconde ou plus) et montrait seulement une faible dépendance à la force appliquée, contrairement à l’ATP pour lequel les temps de dwell augmentent fortement avec la charge. Dans l’ATPγS, la plupart des mouvements arrière étaient des reculs nets d’un pas plutôt que de longs « glissements », ce qui suggère un mode de mouvement sous-jacent différent.
Un état de pause caché mis en lumière
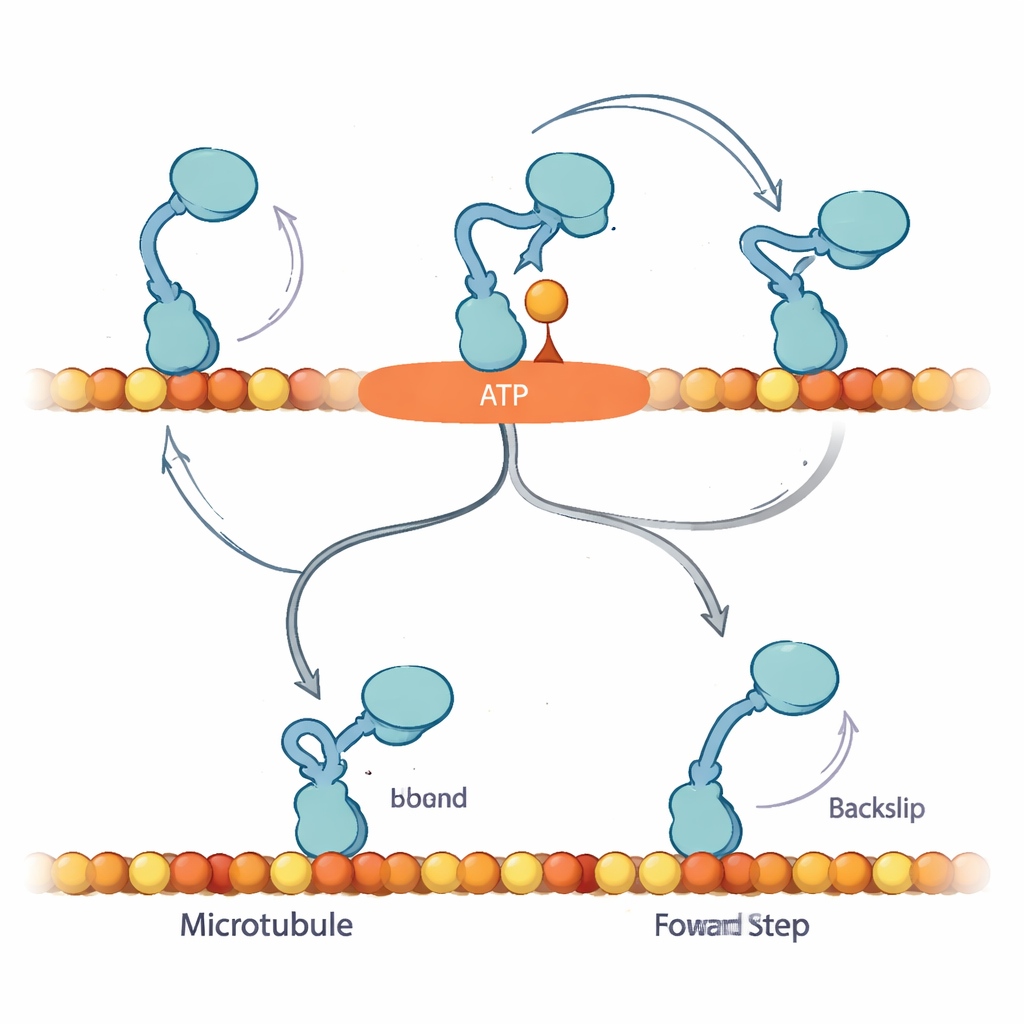
Pour expliquer ces observations, les auteurs proposent que lorsqu’un nucléotide se lie initialement, le kinésine entre dans un état jusqu’alors non reconnu, qu’ils appellent « Await-Isomerisation » (AI). Dans cet état, l’ATP (ou l’ATPγS) est lié et le moteur est prêt à effectuer un pas, mais un segment structurel clé, le linker cervical, n’est pas encore verrouillé et le site actif n’est pas prêt à hydrolyser le carburant. Depuis l’état AI, la tête libre diffuse et peut, en principe, se lier soit en avant soit en arrière sur la piste. Avec l’ATP normal, l’état AI est fugace : il se convertit rapidement en un état « fermé » où le linker cervical se fixe, l’hydrolyse progresse et la tête en suspension est dirigée vers le site avant, favorisant fortement l’avancée. En présence d’abondant ATPγS, cette conversion est ralentie et l’état AI devient surpeuplé, ouvrant une voie annexe dans laquelle le moteur effectue plus souvent de vrais reculs de 8 nanomètres.
Pourquoi une pause contrôlée compte
Les résultats suggèrent que le mécanisme de biais du kinésine dépend de sa capacité à marquer une pause en toute sécurité dans l’état AI sans consommer le carburant, en attendant que le linker cervical se verrouille pour guider la tête libre vers le prochain site de liaison en avant. Sous charge avec l’ATP, l’avancement et l’hydrolyse du carburant sont étroitement couplés : si le moteur n’achève pas un pas avant dans les délais, il peut glisser vers l’arrière, mais cela reste un résultat relativement rare. Avec l’ATPγS, l’état AI prolongé expose une voie cachée de recul, réduisant le biais vers l’avant tout en permettant néanmoins un mouvement net vers l’avant. Pour un lecteur non spécialiste, le message clé est que ce minuscule moteur ne se contente pas de brûler du carburant et d’avancer mécaniquement ; il utilise une salle d’attente intégrée — une pause contrôlée — pour décider d’avancer, de rester en place ou de reculer. La modification subtile du carburant révèle cette logique cachée et montre comment les cellules ajustent finement le mouvement pour maintenir le trafic moléculaire dans la bonne direction, même sous contrainte.
Citation: Karnawat, V., Toleikis, A., Carter, N.J. et al. ATPγS substantially defeats the biasing mechanism for kinesin steps. Nat Commun 17, 2891 (2026). https://doi.org/10.1038/s41467-026-69573-z
Mots-clés: moteur kinésine, moteurs moléculaires, analogues de l'ATP, biophysique monocentrique, transport sur microtubules