Clear Sky Science · sv
ATPγS undergräver i hög grad den biasande mekanismen för kinesinsteg
Hur cellens lastbärare håller kursen
Inuti varje cell transporterar små proteinstegare kallade kinesiner last längs mikroskopiska spår och hjälper till att bygga, reparera och dela cellen. Som lastbilar på en motorväg måste dessa vandrare i huvudsak fortsätta framåt, även när de drar tunga laster. Denna studie ställer en förledande enkel fråga: vad händer med kinesins förmåga att gå framåt om vi något ändrar dess bränsle? Svaret avslöjar ett oväntat paus-och-gå-tillstånd som hjälper motorn att hålla kursen.
En molekylär vandrare och dess sedvanliga bränsle
Kinesin-1 är ett tvåbent motorprotein som vandrar längs mikrotubuli, styva filament som korsar cellen. Varje steg är ungefär 8 nanometer och drivs av den energirika molekylen ATP. Ett huvud håller hårt i spåret medan det andra svänger framåt, och de alternerar i ett hand-över-hand-sätt. Under motriktad kraft—som en last som drar bakåt—finner motorn ändå att framåtrörelse föredras, tack vare en intern ”bias”-mekanism som gör framåtrörelser mer sannolika än bakåtslirtningar.
Byta ATP mot en långsammare dubbelgångare
För att undersöka hur denna bias fungerar bytte forskarna ut ATP mot ATPγS, en nästan identisk molekyl som motorn spjälkar mycket långsammare. Med enkla molekylära optiska fällor—lasertänger som kan hålla i en pärla fäst vid en enskild kinesin—mätte de hur ofta kinesin tog framåtroch bakåtssteg under olika laster och bränsleförhållanden. Vid låga koncentrationer av ATPγS (1 mikromolar) betedde sig motorn mycket som med ATP: den gick processivt framåt, och bakåtgående händelser blev vanligare först när lasten ökade.
När för mycket långsamt bränsle bryter biasen
Bilden förändrades dramatiskt vid hög ATPγS-koncentration (1 millimolar). Kinesin fortsatte att ta steg längs mikrotubuli och nådde liknande stoppkrafter, men korta 8-nanometers baksteg blev mycket vanligare även vid låga laster. Förhållandet mellan fram- och baksteg, som normalt minskar kraftigt när lasten ökar, blev nästan platt: lasten spelade knappt någon roll. Samtidigt var väntetiden före varje steg—dwell-tiden—lång (ungefär en halv sekund eller mer) och visade bara svag beroende av den applicerade kraften, till skillnad från i ATP där dwell-tiderna ökade brant med last. I ATPγS var de flesta bakåtrörelser rena, enkelstegsretreater snarare än långa ”slips”, vilket tyder på en annan underliggande rörelse.
Ett dolt paus-tillstånd träder fram
För att förklara dessa mönster föreslår författarna att när ett nukleotid först binder går kinesin in i ett tidigare oupptäckt ”Await-Isomerisation” (AI)-tillstånd. I detta tillstånd är ATP (eller ATPγS) bundet och motorn är förberedd att ta ett steg, men en nyckelstruktur, neck-linkern, har ännu inte låst sig och den aktiva ytan är inte redo att spjälka bränslet. Från AI-tillståndet diffunderar det fria huvudet och kan i princip binda både framåt och bakåt på spåret. I normalt ATP är AI-tillståndet flyktigt: det konverterar snabbt till ett ”stängt” tillstånd där neck-linkern dockar, hydrolysen fortlöper och det fria huvudet styrs mot det framåtriktade bindningsstället, vilket starkt gynnar framsteg. Vid rikligt med ATPγS fördröjs denna konversion och AI-tillståndet överpopulationeras, vilket öppnar en sidoväg där motorn oftare tar genuina 8-nanometers baksteg.
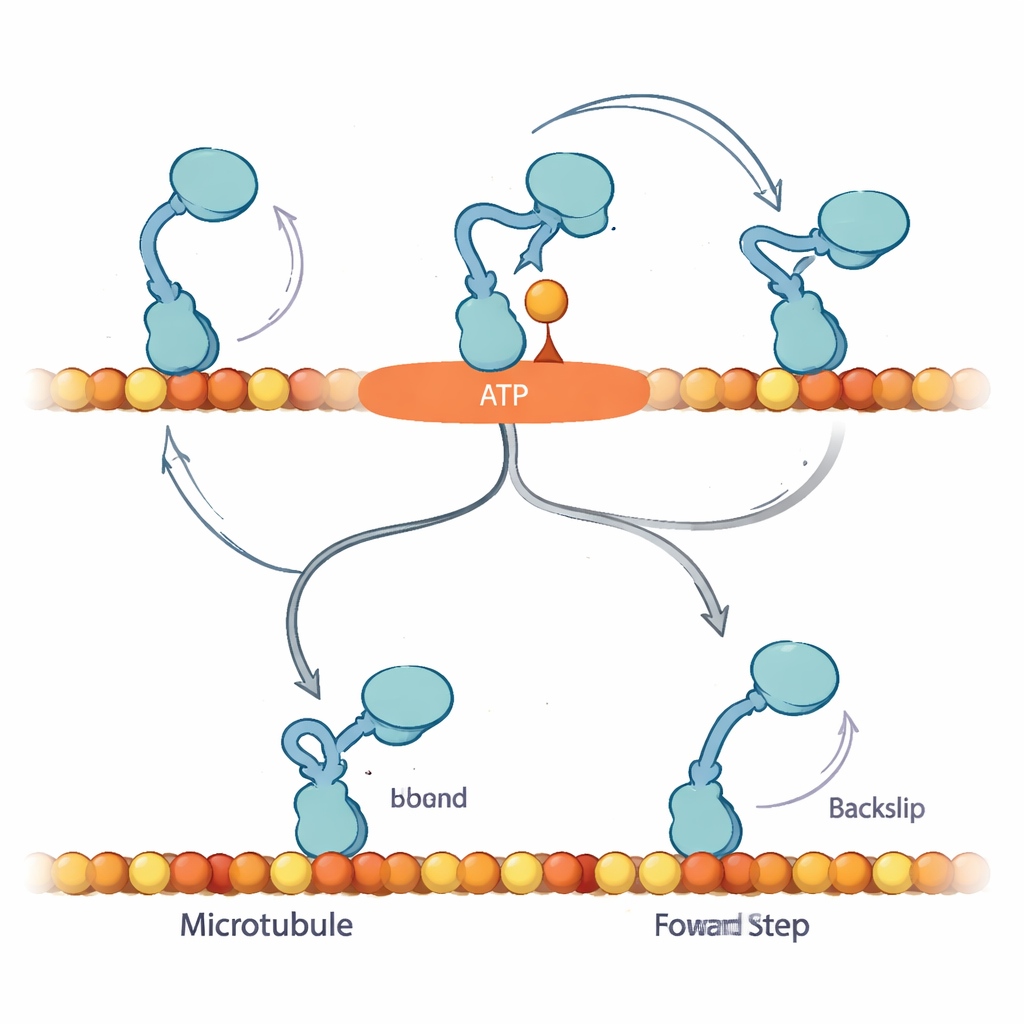
Varför en kontrollerad paus spelar roll
Resultaten tyder på att kinesins bias-mekanism förlitar sig på möjligheten att säkert pausa i AI-tillståndet utan att förbruka bränsle, och vänta på att neck-linker-dockning ska styra det fria huvudet till nästa framåtbundna läge. Under last med ATP är framsteg och bränsleuppluckring tätt kopplade: om motorn misslyckas med att slutföra ett framsteg i tid kan den glida bakåt, men detta är ett relativt sällsynt utfall. I ATPγS exponerar det förlängda AI-tillståndet en dold bakstegsväg, vilket minskar framåtbiasen men fortfarande tillåter nettoframåtrörelse. För en lekmannabetraktare är huvudbudskapet att denna lilla motor inte bara bränner bränsle och marscherar framåt; den använder ett inbyggt väntrum—en kontrollerad paus—för att avgöra om den ska avancera, stanna eller retirera. Att subtilt förändra bränslet avslöjar denna dolda logik och visar hur celler finjusterar rörelsen för att hålla sitt molekylära trafikflöde i rätt riktning, även under belastning.
Citering: Karnawat, V., Toleikis, A., Carter, N.J. et al. ATPγS substantially defeats the biasing mechanism for kinesin steps. Nat Commun 17, 2891 (2026). https://doi.org/10.1038/s41467-026-69573-z
Nyckelord: kinesinmotor, molekylära motorer, ATP-analoger, enskelmolekylsbiologi, mikrotubulitransport