Clear Sky Science · nl
ATPγS ondermijnt grotendeels het biaseringsmechanisme voor kinesin-stappen
Hoe cellulaire vrachtauto’s hun richting behouden
In elke cel slepen piepkleine eiwit"lopers" genaamd kinesines ladingen langs microscopische spoorwegen en helpen zo bij opbouw, reparatie en deling van de cel. Net als vrachtwagens op een snelweg moeten deze lopers grotendeels vooruit blijven bewegen, zelfs wanneer ze zware lasten trekken. Deze studie stelt een bedrieglijk eenvoudige vraag: wat gebeurt er met het vermogen van kinesine om vooruit te lopen als we zijn brandstof lichtjes veranderen? Het antwoord onthult een onverwachte pauze-en-ga-toestand die de motor helpt op koers te blijven.
Een moleculaire wandelaar en zijn gebruikelijke brandstof
Kinesine-1 is een tweebenig motoreiwit dat langs microtubuli loopt, stijve filamenten die door de cel kruisen. Elke stap is ongeveer 8 miljardste van een meter en wordt aangedreven door het energierijke molecuul ATP. Het ene kopje van de kinesine houdt zich stevig aan het spoor vast terwijl het andere naar voren zwaait, en de twee wisselen elkaar af in een hand-over-hand beweging. Onder tegengestelde kracht—zoals een achterwaarts trekkende lading—geeft de motor nog steeds de voorkeur aan voorwaartse stappen, dankzij een intern "biaserings"mechanisme dat voorwaartse beweging waarschijnlijker maakt dan achterwaartse uitglijders.
ATP verwisselen voor een trager gelijkend molecuul
Om te onderzoeken hoe deze bias werkt, vervingen de onderzoekers ATP door ATPγS, een vrijwel identiek molecuul dat de motor veel langzamer afbreekt. Met single-molecule optische vallen—laser-tangetjes die een bolletje aan een enkele kinesine kunnen vasthouden— meetten ze hoe vaak kinesine voor- versus achterwaartse stappen nam onder verschillende belastingen en brandstofcondities. Bij lage concentraties ATPγS (1 micromolair) gedroeg de motor zich grotendeels zoals met ATP: hij liep procesmatig vooruit, en achterwaartse gebeurtenissen werden pas vaker naarmate de belasting toenam.
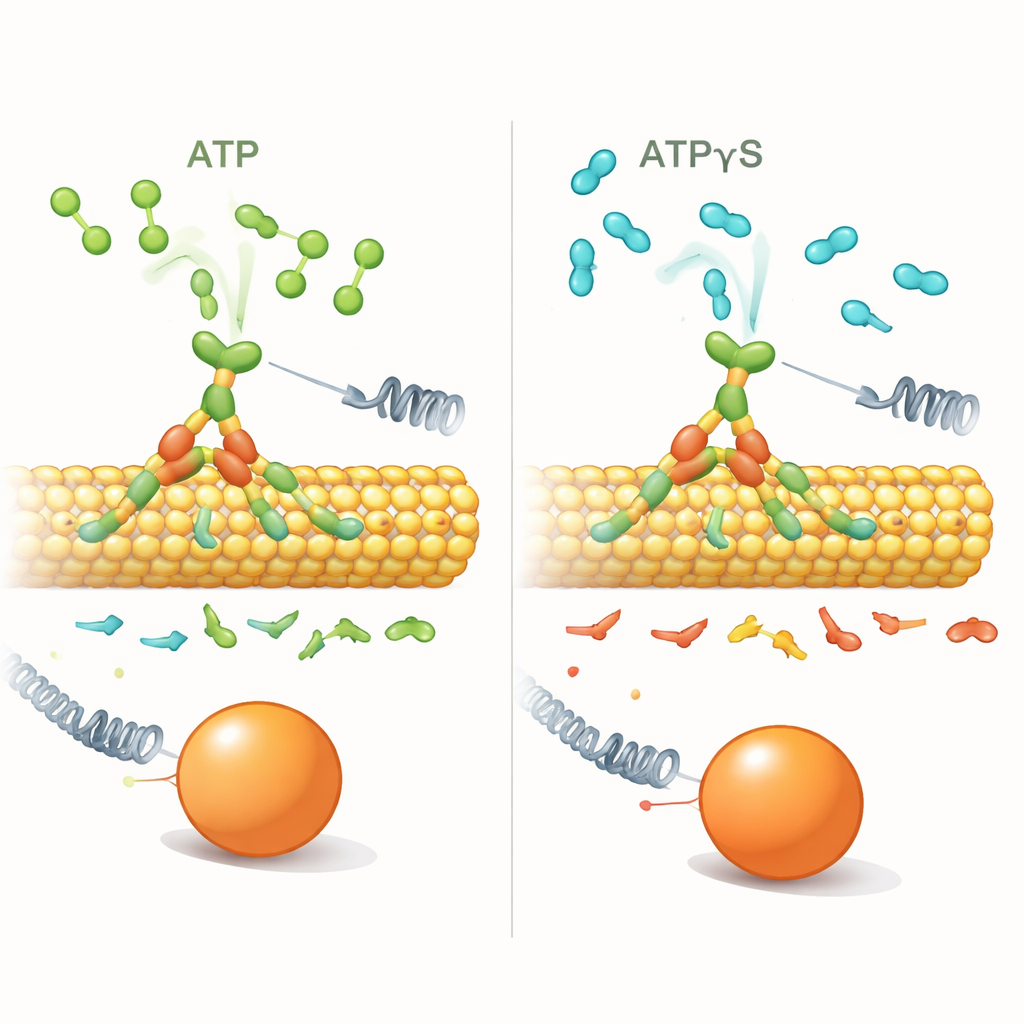
Wanneer te veel trage brandstof de bias doorbreekt
Het beeld veranderde dramatisch bij hoge ATPγS-concentratie (1 millimolair). Kinesine stapte nog steeds langs de microtubulus en bereikte vergelijkbare stallkrachten, maar korte 8-nanometer achterwaartse stappen werden veel frequenter, zelfs bij lage belastingen. De verhouding van voor- tot achterwaartse stappen, die normaal scherp daalt als de belasting toeneemt, werd vrijwel vlak: de belasting deed er nauwelijks meer toe. Tegelijkertijd waren de wachttijden voor elke stap—de dwell—lang (ongeveer een halve seconde of meer) en toonden ze slechts een zwakke afhankelijkheid van de toegepaste kracht, in tegenstelling tot bij ATP waar dwell-tijden sterk toenamen met belasting. In ATPγS waren de meeste achterwaartse bewegingen nette, enkelvoudige terugstappen in plaats van lange "slips", wat duidt op een andere onderliggende beweging.
Een verborgen pauzetoestand komt aan het licht
Om deze patronen te verklaren, stellen de auteurs voor dat wanneer een nucleotide eerst bindt, kinesine in een voorheen niet herkende "Await-Isomerisation" (AI)-toestand komt. In deze toestand is ATP (of ATPγS) gebonden en is de motor klaar om te stappen, maar een cruciaal structureel segment, de neklinker, is nog niet vergrendeld en de actieve plek is nog niet klaar om de brandstof af te breken. Vanuit de AI-toestand kan het vrije kopje diffusief bewegen en in principe naar voren of achteren op het spoor binden. Bij normaal ATP is de AI-toestand vluchtig: hij zet zich snel om in een "gesloten" toestand waarin de neklinker vastklikt, hydrolyse voortgaat en het gebonden kopje naar de voorwaartse plaats wordt gestuurd, waardoor voorwaartse voortgang sterk wordt bevoordeeld. Bij overvloedig ATPγS wordt deze conversie vertraagd en raakt de AI-toestand overbevolkt, waardoor een zijpad opent waarin de motor vaker echte 8-nanometer achterstappen maakt.
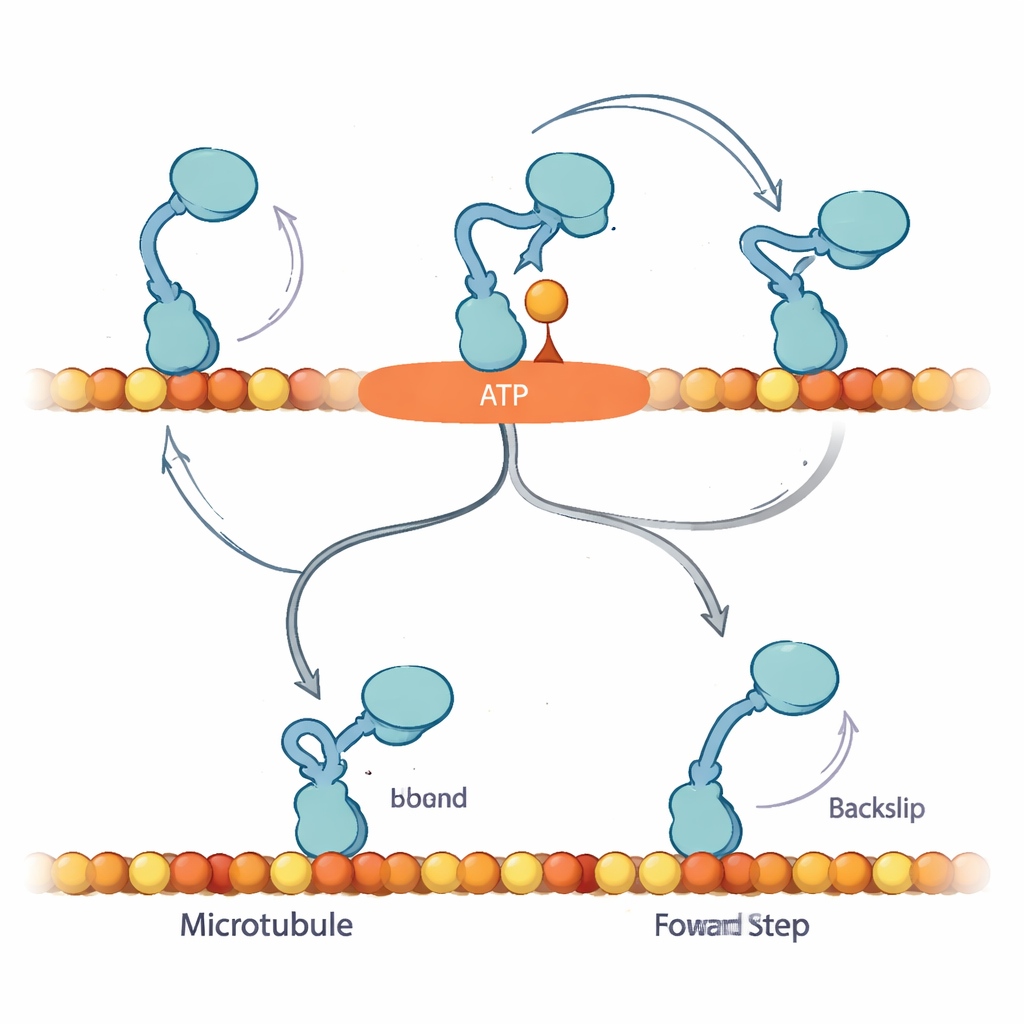
Waarom een gecontroleerde pauze ertoe doet
De bevindingen suggereren dat het biaseringsmechanisme van kinesine afhangt van het vermogen om veilig te pauzeren in de AI-toestand zonder brandstof te verbruiken, wachtend op het vastklikken van de neklinker om het vrije kopje naar de volgende voorwaartse bindingsplaats te leiden. Onder belasting met ATP zijn voorwaarts stappen en brandstofafbraak strak gekoppeld: als de motor er niet in slaagt een voorwaartse stap op tijd te voltooien, kan hij achteruit glijden, maar dat is een relatief zeldzame uitkomst. In ATPγS onthult de verlengde AI-toestand een verborgen terugstaproute, waardoor de voorwaartse bias afneemt maar netto nog steeds voorwaartse beweging mogelijk blijft. Voor een niet-specialistische lezer is de kernboodschap dat deze piepkleine motor niet simpelweg brandstof verbrandt en vooruit marcheert; hij gebruikt een ingebouwde wachtkamer—een gecontroleerde pauze—om te beslissen of hij vooruitgaat, vasthoudt of terugtreedt. Het subtiel veranderen van de brandstof onthult deze verborgen logica en laat zien hoe cellen beweging fijnregelen om hun moleculaire verkeer onder spanning in de juiste richting te houden.
Bronvermelding: Karnawat, V., Toleikis, A., Carter, N.J. et al. ATPγS substantially defeats the biasing mechanism for kinesin steps. Nat Commun 17, 2891 (2026). https://doi.org/10.1038/s41467-026-69573-z
Trefwoorden: kinesine-motor, moleculaire motoren, ATP-analogen, single-molecule biophysica, microtubulustransport