Clear Sky Science · de
ATPγS untergräbt weitgehend den Bias-Mechanismus für Kinesin-Schritte
Wie zelluläre Frachttransporteure ihre Richtung halten
Im Inneren jeder Zelle ziehen winzige Protein‑„Spaziergänger“ namens Kinesine Fracht entlang mikroskopischer Schienen und helfen so beim Aufbau, der Reparatur und der Teilung der Zelle. Wie Lkw auf einer Autobahn müssen diese Wanderer überwiegend vorwärts fahren, selbst wenn sie schwere Lasten ziehen. Diese Studie stellt eine auf den ersten Blick einfache Frage: Was passiert mit der Vorwärtsfähigkeit von Kinesin, wenn man seinen Treibstoff leicht verändert? Die Antwort offenbart einen unerwarteten Pause‑und‑Geh‑Zustand, der dem Motor hilft, auf Kurs zu bleiben.
Ein molekularer Wanderer und sein üblicher Treibstoff
Kinesin‑1 ist ein zweibeiniger Motorprotein, das entlang von Mikrotubuli läuft, starren Filamenten, die die Zelle durchziehen. Jeder Schritt beträgt etwa 8 Milliardstel Meter und wird von dem energiereichen Molekül ATP angetrieben. Ein Kopf des Kinesins hält sich fest am Gleis, während der andere nach vorne schwingt; die beiden wechseln sich in einer Hand‑über‑Hand‑Bewegung ab. Unter Gegenkraft — etwa wenn eine Last nach hinten zieht — bevorzugt der Motor dennoch Vorwärtsschritte, dank eines internen „Bias“-Mechanismus, der Vorwärtsbewegungen wahrscheinlicher macht als Rückrutsche.
ATP gegen ein langsameres Doppelgänger‑Molekül tauschen
Um zu untersuchen, wie dieser Bias funktioniert, ersetzten die Forscher ATP durch ATPγS, ein fast identisches Molekül, das vom Motor deutlich langsamer gespalten wird. Mit Einzelmolekül‑Optikfallen — Laserpinzetten, die eine an ein einzelnes Kinesin gebundene Kugel fixieren können — maßen sie, wie oft Kinesin unter verschiedenen Last‑ und Treibstoffbedingungen vorwärts versus rückwärts schritt. Bei niedrigen Konzentrationen von ATPγS (1 Mikromolar) verhielt sich der Motor sehr ähnlich wie mit ATP: er ging prozessiv vorwärts, und Rückwärtsereignisse wurden erst mit zunehmender Last häufiger.
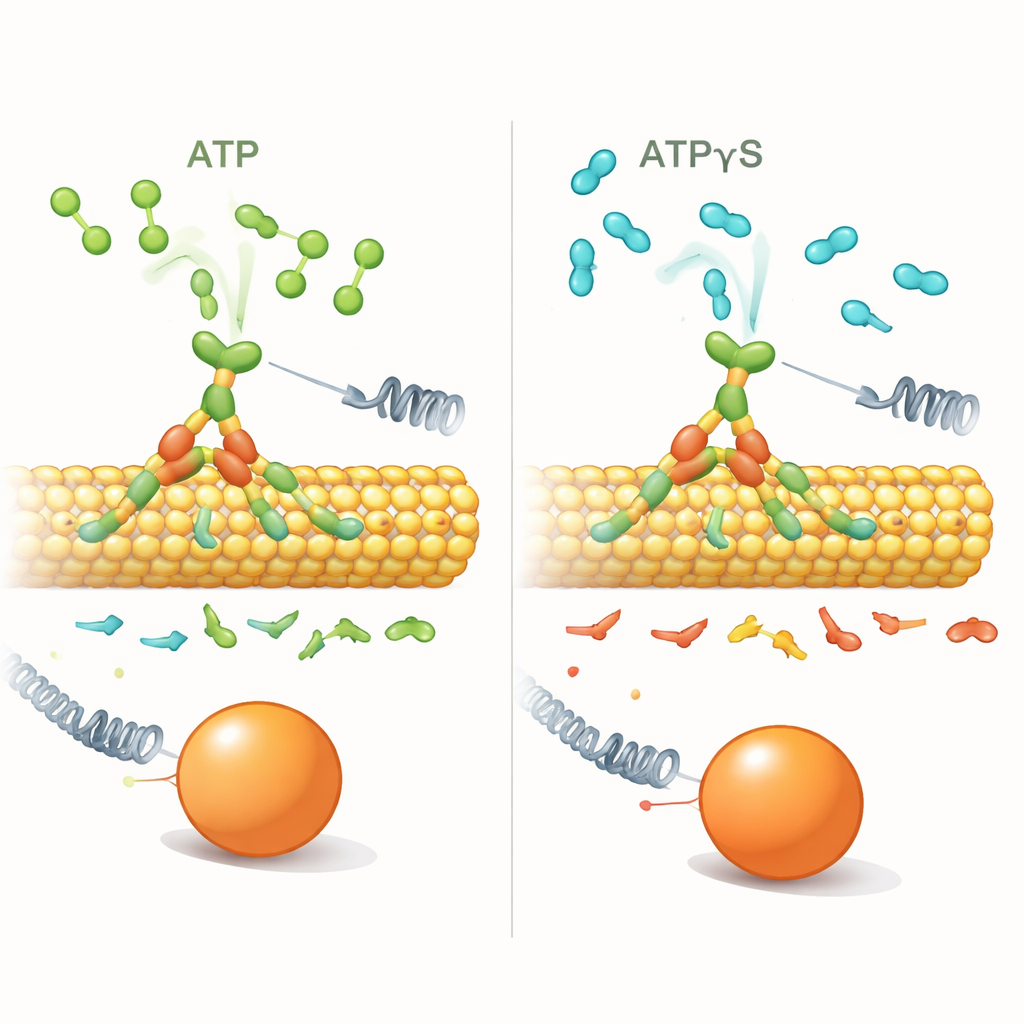
Wenn zu viel langsamer Treibstoff den Bias aufbricht
Das Bild änderte sich dramatisch bei hoher ATPγS‑Konzentration (1 Millimolar). Kinesin schritt weiterhin am Mikrotubulus entlang und erreichte ähnliche Stallkräfte, doch kurze 8‑Nanometer‑Rückschritte wurden nun selbst bei geringen Lasten deutlich häufiger. Das Verhältnis von Vorwärts‑ zu Rückwärtsschritten, das normalerweise mit zunehmender Last stark abfällt, wurde nahezu flach: die Last spielte kaum noch eine Rolle. Gleichzeitig waren die Wartezeiten vor jedem Schritt — die Dwell‑Times — lang (etwa eine halbe Sekunde oder mehr) und zeigten nur eine schwache Abhängigkeit von der angelegten Kraft, anders als bei ATP, wo die Dwell‑Times mit der Last stark anstiegen. In ATPγS waren die meisten Rückschritte saubere Einzelschritt‑Retreats statt langer „Slips“, was auf eine andere zugrundeliegende Bewegung hindeutet.
Ein verborgener Pausenzustand tritt zutage
Um diese Muster zu erklären, schlagen die Autoren vor, dass beim ersten Binden eines Nukleotids Kinesin in einen bislang unerkannte n „Await‑Isomerisation“‑(AI‑)Zustand gelangt. In diesem Zustand ist ATP (oder ATPγS) gebunden und der Motor zum Schritt vorbereitet, aber ein wichtiger Strukturabschnitt, der Neck‑Linker, hat sich noch nicht verriegelt und das aktive Zentrum ist noch nicht bereit, den Treibstoff zu spalten. Aus dem AI‑Zustand kann der freie Kopf diffundieren und theoretisch entweder vorwärts oder rückwärts an das Gleis binden. Bei normalem ATP ist der AI‑Zustand kurzlebig: er wandelt sich schnell in einen „geschlossenen“ Zustand um, in dem der Neck‑Linker andockt, Hydrolyse abläuft und der freie Kopf zum vorderen Bindungsort gesteuert wird, wodurch Vorwärtsbewegung stark begünstigt wird. Bei reichlich ATPγS wird diese Umwandlung verlangsamt und der AI‑Zustand überbevölkert, wodurch ein Nebenweg geöffnet wird, in dem der Motor häufiger echte 8‑Nanometer‑Rückschritte macht.
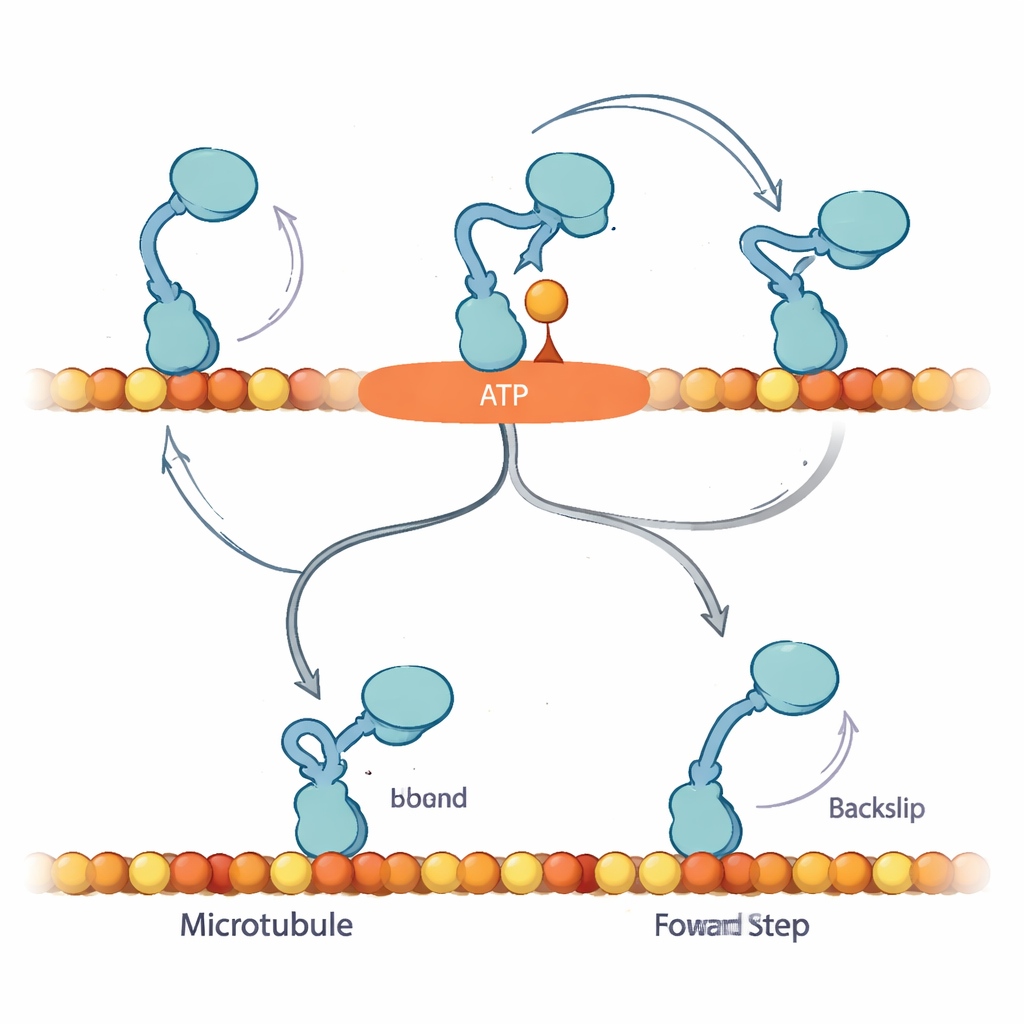
Warum eine kontrollierte Pause wichtig ist
Die Ergebnisse deuten darauf hin, dass Kinesins Bias‑Mechanismus auf der Fähigkeit beruht, sicher im AI‑Zustand zu pausieren, ohne Treibstoff zu verbrauchen, und auf die Dockung des Neck‑Linkers zu warten, die den freien Kopf zum nächsten vorderen Bindungsplatz leitet. Unter Last und mit ATP sind Vorwärtsschritte und Treibstoffabbau eng gekoppelt: Gelingt dem Motor ein Vorwärtsschritt nicht rechtzeitig, kann er rückwärts rutschen, doch das ist vergleichsweise selten. In ATPγS macht der verlängerte AI‑Zustand einen verborgenen Rückschrittweg sichtbar, reduziert die Vorwärtsbias, erlaubt aber dennoch netto Vorwärtsbewegung. Für eine allgemeine Leserschaft ist die Kernbotschaft, dass dieser winzige Motor nicht einfach nur Treibstoff verbrennt und vorwärts marschiert; er nutzt einen eingebauten Wartebereich — eine kontrollierte Pause — um zu entscheiden, ob er voranschreitet, verharrt oder zurückweicht. Eine subtile Veränderung des Treibstoffs legt diese versteckte Logik offen und zeigt, wie Zellen die Bewegung feinjustieren, damit ihr molekularer Verkehr selbst unter Belastung in die richtige Richtung fließt.
Zitation: Karnawat, V., Toleikis, A., Carter, N.J. et al. ATPγS substantially defeats the biasing mechanism for kinesin steps. Nat Commun 17, 2891 (2026). https://doi.org/10.1038/s41467-026-69573-z
Schlüsselwörter: Kinesin‑Motor, molekulare Motoren, ATP‑Analoga, Einzelmolekül‑Biophysik, Mikrotubulus‑Transport