Clear Sky Science · tr
İnsan TREX-2 kompleksi tarafından mRNA ihracatının düzenlenmesine ilişkin moleküler içgörüler
Hücreler hangi mesajların çekirdekten çıkmasına izin vereceğine nasıl karar veriyor
Her saniye, hücrelerimiz diğer hücre bileşenlerine hangi proteinlerin üretileceğini söyleyen binlerce genetik "mesajı" haberci RNA (mRNA) biçiminde gönderir. Yanlış mesajların geçmesine izin vermek—veya onları yanlış zamanda salmak—gelişim bozukluklarından kansere kadar ciddi problemlere yol açabilir. Bu çalışma, nükleer porlardaki anahtar bir kapı bekçisi kompleksi olan TREX-2’nin, bir moleküler motor protein olan UAP56 ile birlikte mRNA paketinin ne zaman çekirdeği terk etmeye hazır olduğunu nasıl karar verdiğini ortaya koyuyor.
Gevşek taslaktan mühürlenmiş mesaja
Bir mRNA çekirdeği terk etmeden önce dikkatle işlenmeli ve kararlı, ihracata hazır bir parçacık oluşturmak için birçok proteinle sarılmalıdır. Önceki çalışmalar, TREX adı verilen bir montaj hattının mRNA’yı paketlediğini ve hücresel yakıt (ATP) kullanan ve RNA’ya sıkı bağlanan motor benzeri bir protein olan UAP56 ile birlikte kılavuzladığını göstermişti. TREX-2 adlı ikinci bir kompleks ise nükleer zardaki kapılar olan nükleer porlara bağlı olarak oturur ve ihracat için elzem olduğu bilinmektedir. Ancak TREX-2’nin UAP56’e bağlı mRNA’yı nasıl tanıdığı ve mesajın son ihracat taşıyıcısına nasıl aktarıldığı belirsizliğini koruyordu. 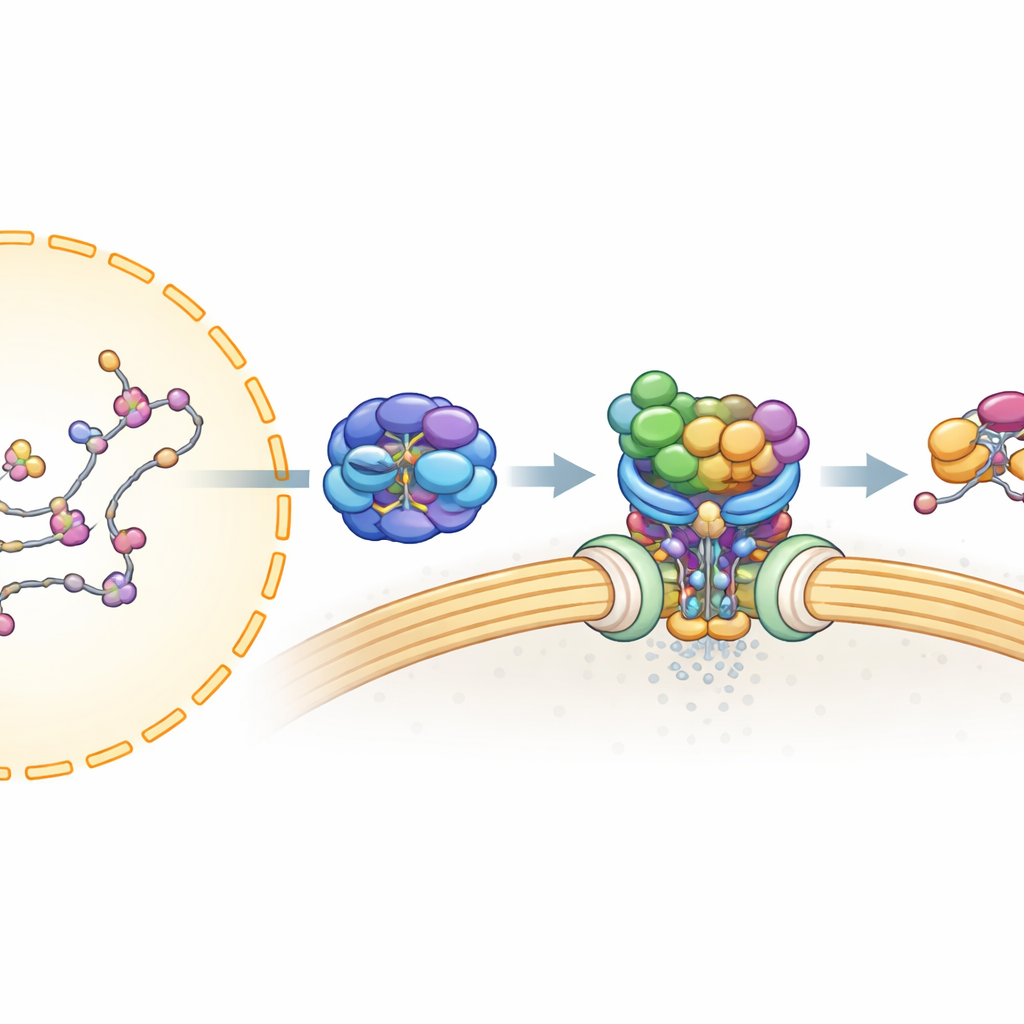
Nükleer kapıda moleküler bir tokalaşma
Yazarlar, TREX-2 ile fiziksel olarak hangi proteinlerin ilişki kurduğunu haritalamak için insan hücreleri, protein saflaştırma ve kütle spektrometrisi kullandılar. TREX bileşenlerinin çoğu değil, UAP56’nın TREX-2’nin çekirdek iskeletine—GANP, PCID2 ve DSS1 adlı proteinlerden oluşan yapıya—güçlü bağlandığını buldular. Bu bulgu, paketlenmiş mRNA’yı TREX-2’ye getiren özel bir adaptör olarak UAP56’yı işaret etti. İlerleyen deneyler, UAP56 ile TREX-2’nin yalnızca UAP56 bir nükleotit (ATP veya ADP gibi bir yakıt ilişkili molekül) tuttuğunda sıkı bir kompleks oluşturduğunu gösterdi; bu da TREX-2’nin ihracat döngüsü sırasında UAP56’nın belirli bir "yüklü" durumunu tanıdığını düşündürüyordu.
Kapı bekçisini atomik ayrıntıda görmek
Bu etkileşimi yüksek çözünürlükte anlamak için ekip, molekülleri şokla dondurarak görüntüleyen ve araştırmacıların 3B şekillerini yeniden inşa etmelerine olanak veren kriyo-elektron mikroskobisine başvurdu. TREX-2’nin tek başına ve UAP56’ye bağlı yapılarını çözdüler. TREX-2’nin çekirdeği V biçimli bir yuva oluşturuyor; GANP ve PCID2 kolları yaparken DSS1 bir tarafta yuvalanıyor. UAP56 bu yuvada bir sapan içindeki top gibi duruyor ve TREX-2 ile esas olarak başlangıçtaki iki esnek döngü (N-terminal bölge) aracılığıyla temas kuruyor. Bu döngüler GANP ve PCID2 üzerindeki pozitif yüklü bir oluğa uzanarak UAP56’yı V biçimli yüzeye kilitliyor. Bu döngülerdeki anahtar amino asitleri değiştirmek—etkili bir şekilde kanca uçlarını kesmek veya körleştirmek—bağlanmayı büyük ölçüde zayıflattı veya ortadan kaldırdı; bu da UAP56’nın bu küçük bölgesinin TREX-2 için ana demirleme noktası olduğunu doğruladı.
TREX-2 motoru nasıl bırakmaya zorlar
Şaşırtıcı bir şekilde, TREX-2 bağlandığında UAP56 RNA’yı tutmayan "açık" bir pozda yakalanmıştır; oysa kompleks RNA varlığında monte edilmişti. Yapı bunun nedenini ortaya koyuyor: GANP’dan gelen kısa bir döngü, etkinleştirici döngü adı verilen yapı, normalde RNA zincirini kavrayan UAP56’nın iki lobu arasına kama gibi giriyor. Bu döngü ayrıca UAP56’nın ATP bağlama yuvasına uzanarak ATP’yi algılayan anahtar yan zincirleri iteliyor. Biyokimyasal analizler TREX-2’nin, etkinleştirici döngü ve N-terminal demirleme döngüleri sağlam olduğunda UAP56’nın ATPaz aktivitesini yaklaşık on kat artırdığını gösterdi. Jel tabanlı RNA serbest bırakma deneylerinde TREX-2, RNA’yı aktif şekilde UAP56’dan sıyırırken, etkinleştirici döngüde tek bir kalıntısı değiştirilmiş bir TREX-2 mutantı çok daha zayıf bir etki gösterdi ve demirleme segmenti olmayan bir UAP56 mutantı hiç yanıt vermedi. 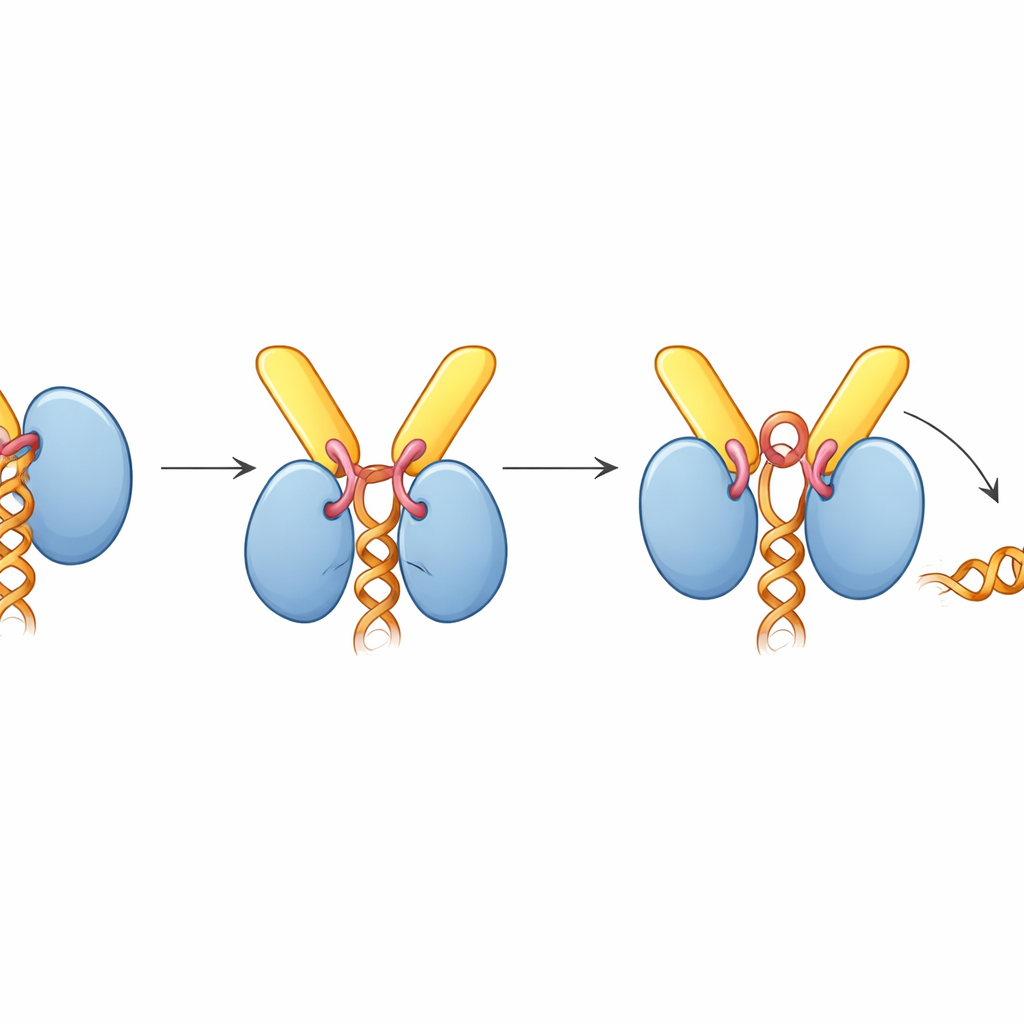
Genetik yükün koordineli devri
Bu bulguları bir araya getirerek yazarlar, mRNA ihracatı için birleşik bir model öneriyor. TREX ve UAP56 önce çekirdek içinde mRNA’yı paketleyip sıkıştırılmış parçacıklar halinde kılıflarlar. Bu parçacıklar sonra UAP56 "sapı"nın nükleer porlarda TREX-2 tarafından yakalanmasına kadar difüz eder. TREX-2’nin etkinleştirici döngüsü UAP56’yı ATP yakıp RNA tutuşunu açan bir konformasyona iterek mRNA’yı serbest bırakır. Serbest kalan mesaj hemen TREX-2 tarafından da işe alınan son ihracat reseptörü (NXF1–NXT1) tarafından yakalanır ve ardından por üzerinden sitoplazmaya taşınır. Basitçe söylemek gerekirse, TREX-2 gelen yükü tanıyan ve onu refakatçisinden yakıt güdümlü bir mekanizma ile çözen ve giden taşıyıcıya aktaran akıllı bir demirleme istasyonu görevi görür.
Bu mikroskobik koreografinin önemi
Bu çalışma, insan hücrelerinin hangi mRNA mesajlarının çekirdekten kaçacağını kontrol etmek için korunmuş bir moleküler anahtarı nasıl kullandığını hemen hemen atomik ayrıntıda açıklıyor. UAP56, durumunun TREX-2 tarafından okunup sıfırlandığı merkezi bir trafik kontrolörü olarak ortaya çıkıyor; bu, yalnızca düzgün paketlenmiş mRNA’nın ihracat makinesine teslim edilmesini sağlar. Bu yolaktaki hatalar genom kararsızlığı ve hastalıklarla ilişkilendirildiğinden, bu koreografinin anlaşılması bu faktörlerdeki mutasyonların nasıl hastalığa neden olabileceğini araştırmak için bir temel sağlar ve uzun vadede mRNA ihracatını terapilerde seçici olarak ayarlamaya yönelik yaklaşımların tasarlanmasına yardımcı olabilir.
Atıf: Gong, X., Tao, R., Ge, X. et al. Molecular insights into mRNA export regulation by the human TREX-2 complex. Nat Commun 17, 3244 (2026). https://doi.org/10.1038/s41467-026-70088-w
Anahtar kelimeler: mRNA ihracatı, TREX-2 kompleksi, UAP56 helikazı, nükleer por, RNA işlenmesi