Clear Sky Science · es
Perspectivas moleculares sobre la regulación de la exportación de ARNm por el complejo TREX-2 humano
Cómo las células deciden qué mensajes pueden salir del núcleo
Cada segundo, nuestras células envían miles de “mensajes” genéticos en forma de ARN mensajero (ARNm) que indican al resto de la célula qué proteínas fabricar. Dejar pasar mensajes incorrectos —o liberarlos en el momento inadecuado— puede causar problemas graves, desde defectos del desarrollo hasta cáncer. Este estudio revela cómo un complejo clave actuando como centinela en el poro nuclear, llamado TREX-2, colabora con una proteína motora molecular, UAP56, para decidir cuándo un paquete de ARNm está listo para abandonar el núcleo.
Del guion suelto al mensaje sellado
Antes de que un ARNm pueda salir del núcleo, debe procesarse cuidadosamente y cubrirse con numerosas proteínas para formar una partícula estable y lista para exportarse. Trabajos anteriores mostraron que una línea de ensamblaje, conocida como TREX, empaqueta el ARNm y lo sujeta junto con UAP56, una proteína con comportamiento de motor que utiliza combustible celular (ATP) y se une fuertemente al ARN. Un segundo complejo, TREX-2, está anclado en los poros nucleares —las puertas en la membrana nuclear— y es esencial para la exportación. Sin embargo, no estaba claro cómo TREX-2 reconoce el ARNm unido a UAP56, ni cómo se transfiere el mensaje al transportador final de exportación. 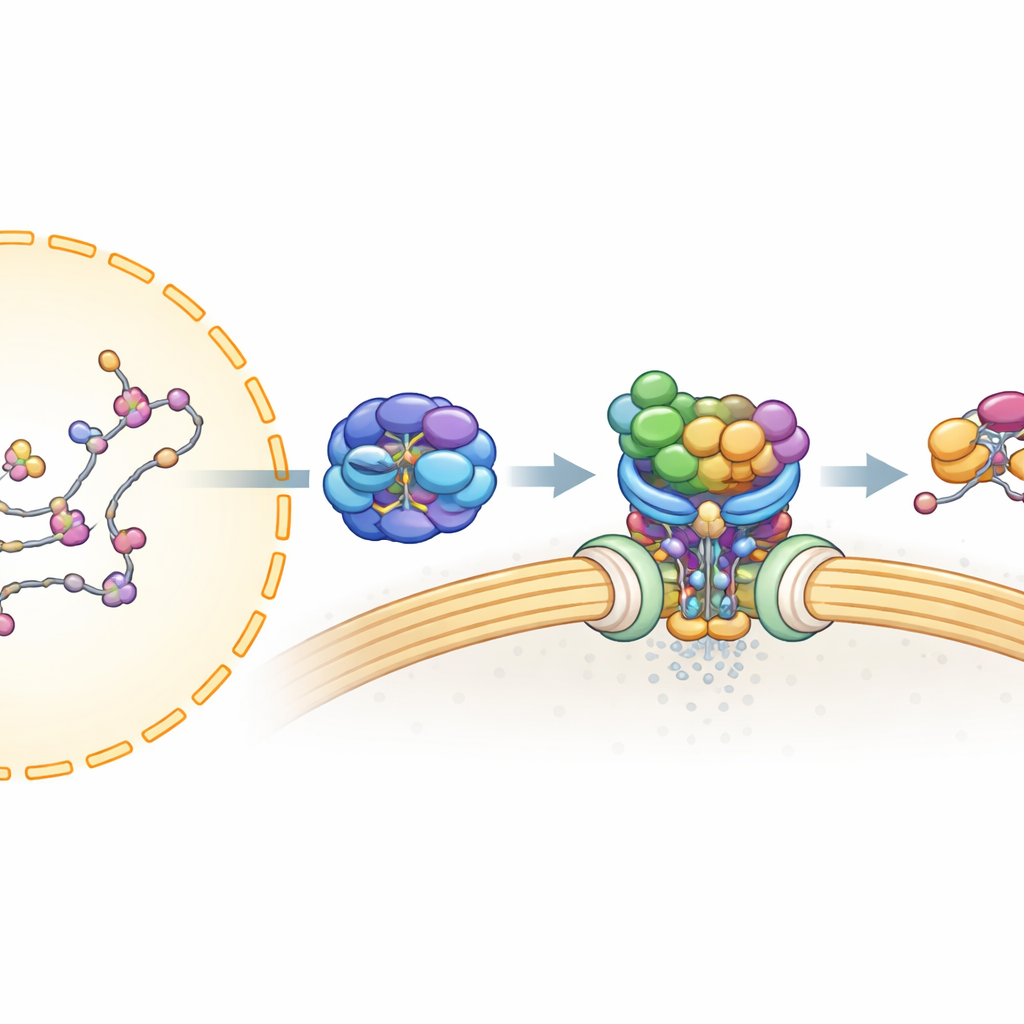
Un apretón de manos molecular en la puerta nuclear
Los autores utilizaron células humanas, purificación de proteínas y espectrometría de masas para mapear qué proteínas se asocian físicamente con TREX-2. Encontraron que UAP56, pero no la mayoría de los demás componentes de TREX, se une fuertemente al andamiaje central de TREX-2, formado por proteínas llamadas GANP, PCID2 y DSS1. Esto señalaba a UAP56 como un adaptador dedicado que trae el ARNm empaquetado a TREX-2. Experimentos posteriores mostraron que UAP56 y TREX-2 solo forman un complejo estable cuando UAP56 contiene un nucleótido (una molécula relacionada con el combustible, como ATP o ADP), lo que sugiere que TREX-2 reconoce un estado «cargado» específico de UAP56 durante el ciclo de exportación.
Ver al centinela con detalle atómico
Para entender esta interacción a alta resolución, el equipo recurrió a la criomicroscopía electrónica, que imagina moléculas flash-congeladas y permite reconstruir sus formas 3D. Resolverion estructuras de TREX-2 solo y unido a UAP56. El núcleo de TREX-2 forma una cuna en forma de V, con GANP y PCID2 formando los brazos y DSS1 alojándose a lo largo de un lado. UAP56 se sitúa en esa cuna como una bola en una honda, contactando a TREX-2 principalmente a través de dos bucles flexibles en su extremo inicial (su región N-terminal). Estos bucles se introducen en una ranura cargada positivamente en GANP y PCID2, fijando UAP56 sobre la superficie en V. Mutar aminoácidos clave en esos bucles —en efecto recortando o embotando las puntas de los ganchos— debilitó o abolió considerablemente la unión, confirmando que esta pequeña región de UAP56 es el principal sitio de anclaje para TREX-2.
Cómo TREX-2 obliga al motor a soltar
Sorprendentemente, cuando TREX-2 está unido, UAP56 queda atrapado en una postura «abierta» que no sujeta ARN, pese a que el complejo se ensambló en presencia de ARN. La estructura revela por qué: un bucle corto de GANP, apodado el bucle activador, se inserta entre los dos lóbulos de UAP56 que normalmente aprietan la hebra de ARN. Ese bucle también alcanza el bolsillo de unión al ATP de UAP56, empujando cadenas laterales clave que normalmente detectan ATP. Ensayos bioquímicos mostraron que TREX-2 aumenta en gran medida la actividad ATPasa de UAP56 —la velocidad a la que consume ATP— aproximadamente diez veces, pero solo cuando el bucle activador y los bucles de anclaje N-terminal están intactos. En experimentos de liberación de ARN basados en gel, TREX-2 arrancó activamente el ARN de UAP56, mientras que un TREX-2 mutante con un solo residuo alterado en el bucle activador tuvo un efecto mucho más débil, y un UAP56 mutante carente de su segmento de anclaje no respondió en absoluto. 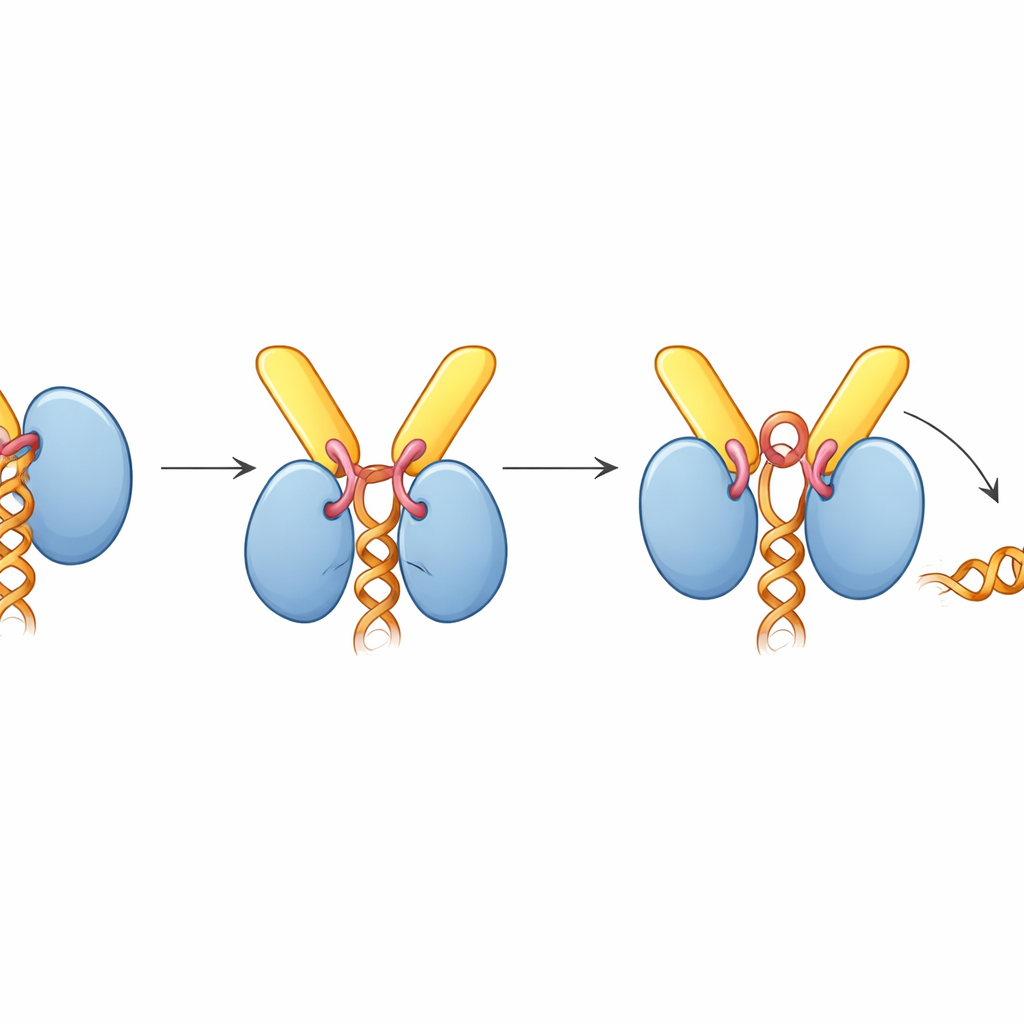
Un traspaso coordinado de la carga genética
Poniendo estos hallazgos en conjunto, los autores proponen un modelo unificado para la exportación de ARNm. TREX y UAP56 primero empaquetan y sujetan el ARNm en partículas compactas dentro del núcleo. Estas partículas difunden hasta que el «asa» de UAP56 es capturada por TREX-2 en el poro nuclear. El bucle activador de TREX-2 empuja a UAP56 hacia una conformación que consume ATP y abre su agarre sobre el ARN, liberando el ARNm. El mensaje liberado es inmediatamente capturado por el receptor final de exportación (NXF1–NXT1), también reclutado por TREX-2, y luego transportado a través del poro hacia el citoplasma. En términos sencillos, TREX-2 actúa como una estación de acoplamiento inteligente que reconoce la carga entrante y desencadena un mecanismo impulsado por combustible para desengancharla de su escolta y pasarla al transportador de salida.
Por qué importa esta coreografía microscópica
Este trabajo explica, con detalle casi atómico, cómo las células humanas usan un interruptor molecular conservado para controlar qué mensajes de ARNm escapan del núcleo. UAP56 emerge como un controlador central del tráfico cuyo estado es leído y restablecido por TREX-2, asegurando que solo el ARNm correctamente empaquetado se entregue a la maquinaria de exportación. Dado que los errores en esta vía se asocian con inestabilidad genómica y enfermedad, comprender esta coreografía proporciona una base para explorar cómo las mutaciones en estos factores podrían causar patologías y, a largo plazo, para diseñar maneras de ajustar selectivamente la exportación de ARNm en terapias.
Cita: Gong, X., Tao, R., Ge, X. et al. Molecular insights into mRNA export regulation by the human TREX-2 complex. Nat Commun 17, 3244 (2026). https://doi.org/10.1038/s41467-026-70088-w
Palabras clave: exportación de ARNm, complejo TREX-2, helicasa UAP56, poro nuclear, procesamiento de ARN