Clear Sky Science · pl
Molukularne wglądy w regulację eksportu mRNA przez ludzki kompleks TREX-2
Jak komórki decydują, które komunikaty mogą opuścić jądro
Co sekundę nasze komórki wysyłają tysiące genetycznych „komunikatów” w postaci informacyjnego RNA (mRNA), które mówią reszcie komórki, jakie białka produkować. Przepuszczenie niewłaściwych komunikatów — lub wypuszczenie ich w nieodpowiednim czasie — może prowadzić do poważnych problemów, od wad rozwojowych po nowotwory. To badanie odsłania, jak kluczowy kompleks kontrolny przy porach jądrowych, zwany TREX-2, współpracuje z białkiem-motorem UAP56, aby zdecydować, kiedy pakiet mRNA jest gotowy do opuszczenia jądra.
Z luźnego zapisu do zapieczętowanej wiadomości
Zanim mRNA będzie mogło opuścić jądro, musi zostać starannie przetworzone i oplecione wieloma białkami, tworząc stabilną cząsteczkę gotową do eksportu. Wcześniejsze prace wykazały, że jedna linia montażowa, znana jako TREX, pakuje mRNA i zaciska je z UAP56 — białkiem przypominającym motor, które wykorzystuje komórkowe paliwo (ATP) i silnie wiąże RNA. Drugi kompleks, TREX-2, jest zakotwiczony przy porach jądrowych — bramach w błonie jądrowej — i wiadomo, że jest niezbędny do eksportu. Jednak w jaki sposób TREX-2 rozpoznaje mRNA związane z UAP56 i jak komunikat jest przekazywany końcowemu nosicielowi eksportowemu, pozostawało niejasne. 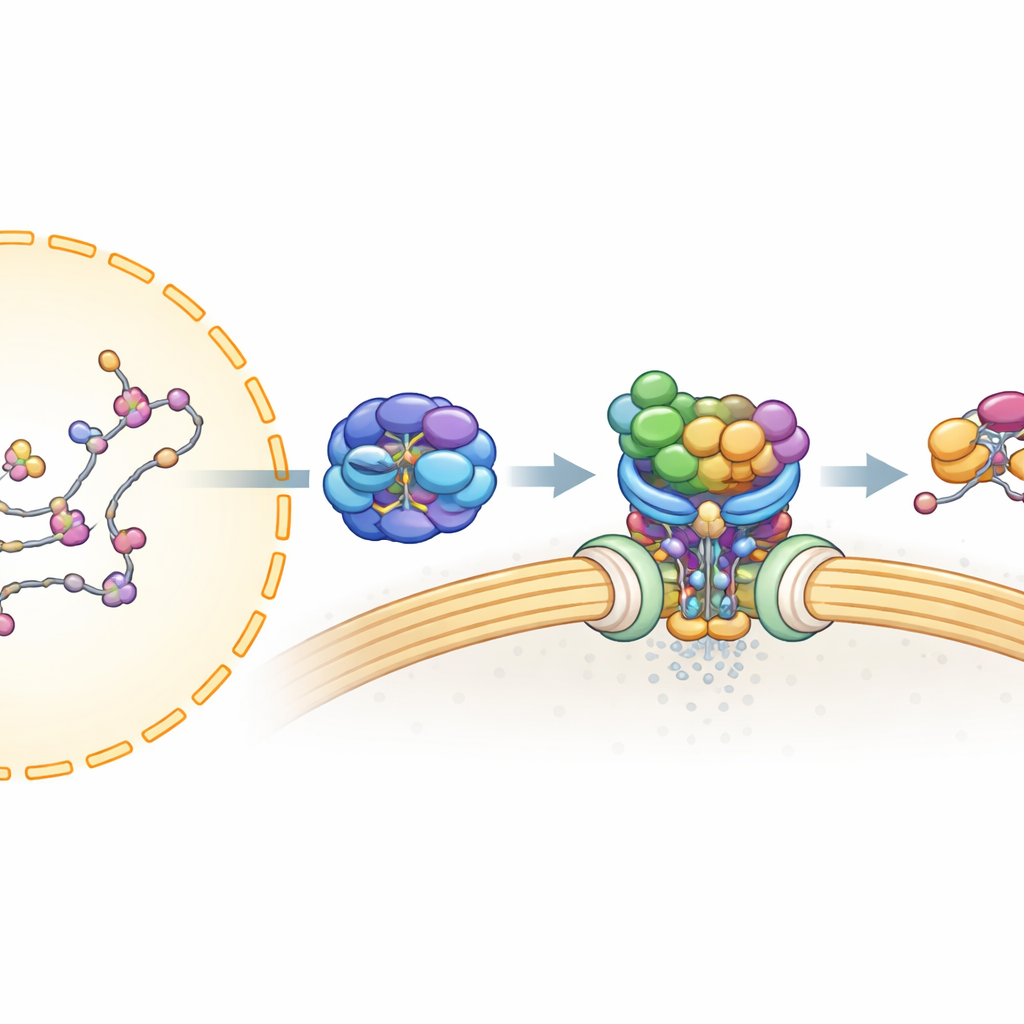
Molekularne uściśnięcie dłoni przy bramie jądrowej
Autorzy użyli komórek ludzkich, oczyszczania białek i spektrometrii masowej, aby zmapować, które białka fizycznie współdziałają z TREX-2. Odkryli, że UAP56, w przeciwieństwie do większości innych składników TREX, silnie wiąże się z rdzeniem rusztowania TREX-2, zbudowanym z białek zwanych GANP, PCID2 i DSS1. Wskazywało to na UAP56 jako wyspecjalizowany adaptor, który przynosi zapakowane mRNA do TREX-2. Dalsze eksperymenty pokazały, że UAP56 i TREX-2 tworzą ścisły kompleks tylko wtedy, gdy UAP56 trzyma nukleotyd (cząsteczkę związaną z paliwem, taką jak ATP lub ADP), co sugeruje, że TREX-2 rozpoznaje specyficzny „zładowany” stan UAP56 w przebiegu cyklu eksportu.
Widok strażnika bramy w detalach atomowych
Aby zrozumieć tę interakcję w wysokiej rozdzielczości, zespół zwrócił się ku krioelektronowej mikroskopii, która obrazując błyskawicznie zamrożone cząsteczki pozwala badaczom odtworzyć ich trójwymiarowe kształty. Opracowali struktury TREX-2 samego oraz w związku z UAP56. Rdzeń TREX-2 tworzy V-kształtną kołyskę, z GANP i PCID2 tworzącymi ramiona, a DSS1 osadzonym wzdłuż jednego boku. UAP56 siedzi w tej kołysce jak kula w temblaku, kontaktując się z TREX-2 głównie przez dwie elastyczne pętle na swoim skrajnym początku (region N-terminalny). Te pętle sięgają do dodatnio naładowanego rowka na GANP i PCID2, blokując UAP56 na powierzchni w kształcie litery V. Mutacje kluczowych aminokwasów w tych pętlach — w istocie przycinające lub tępiące końcówki haczyków — znacznie osłabiły lub zniosły wiązanie, potwierdzając, że ten niewielki region UAP56 jest głównym miejscem dokowania dla TREX-2.
Jak TREX-2 zmusza motor do puszczenia
Zaskakująco, gdy TREX-2 jest związany, UAP56 zostaje uchwycony w „otwartej” pozycji, która nie utrzymuje RNA, mimo że kompleks został zmontowany w obecności RNA. Struktura wyjaśnia dlaczego: krótka pętla z GANP, nazwana pętlą aktywującą, wsuwa się między dwie płaty UAP56, które normalnie zaciskają nici RNA. Ta pętla sięga też do kieszeni wiążącej ATP w UAP56, popychając kluczowe łańcuchy boczne, które zwykle wyczuwają ATP. Analizy biochemiczne wykazały, że TREX-2 znacząco zwiększa aktywność ATP-azy UAP56 — szybkość spalania ATP — około dziesięciokrotnie, lecz tylko wtedy, gdy pętla aktywująca i N-terminalne pętle dokujące są nienaruszone. W żelowych eksperymentach uwalniania RNA TREX-2 aktywnie zdzierał RNA z UAP56, podczas gdy mutant TREX-2 z pojedynczą zmienioną resztą w pętli aktywującej miał znacznie słabszy efekt, a mutant UAP56 pozbawiony segmentu dokującego nie reagował w ogóle. 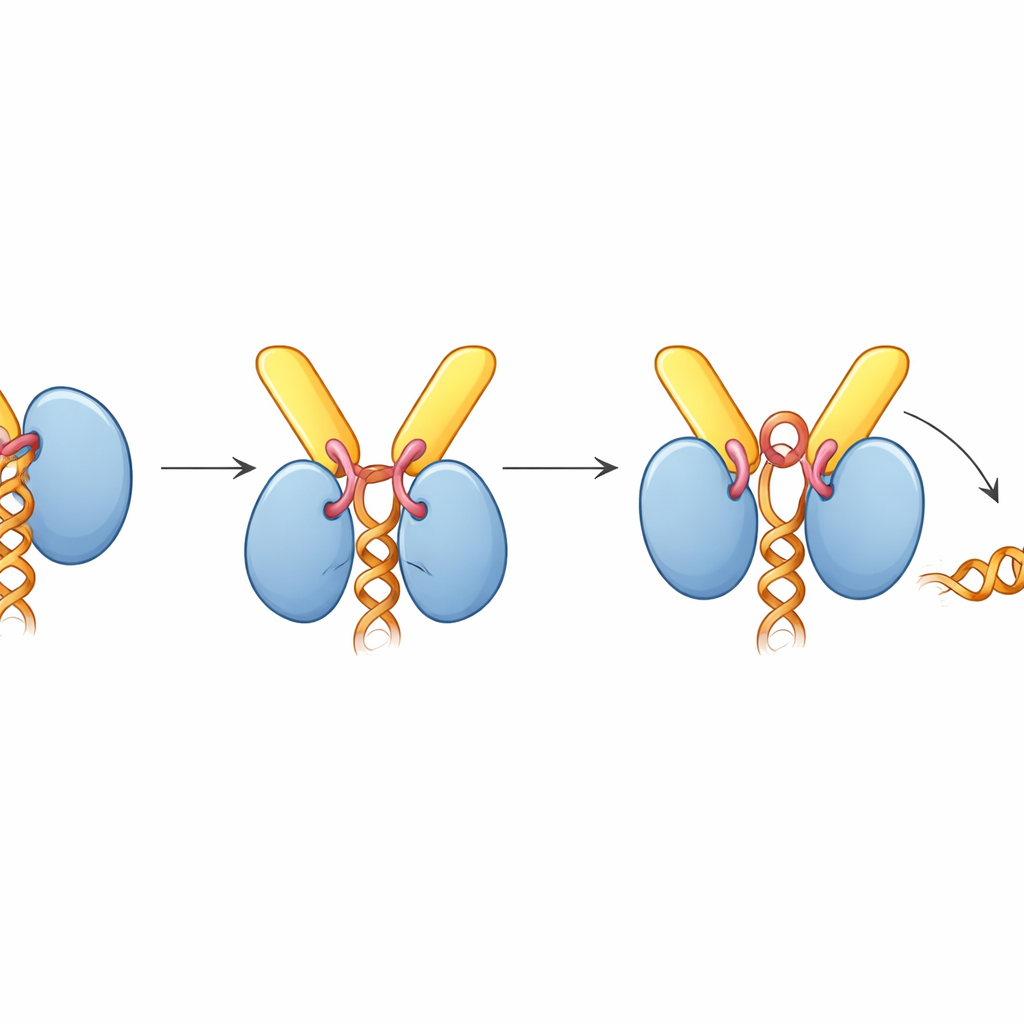
Skoordynowane przekazanie ładunku genetycznego
Skladując te odkrycia, autorzy proponują zunifikowany model eksportu mRNA. TREX i UAP56 najpierw pakują i zaciskają mRNA w zwarte cząsteczki wewnątrz jądra. Te cząsteczki następnie dyfundują, aż „uchwyt” UAP56 zostanie pochwycony przez TREX-2 przy porze jądrowej. Pętla aktywująca TREX-2 wypycha UAP56 w konformację, która spala ATP i rozsuwa jego uchwyt na RNA, uwalniając mRNA. Uwolniona wiadomość jest natychmiast łapana przez końcowy receptor eksportowy (NXF1–NXT1), który również jest rekrutowany przez TREX-2, a następnie transportowana przez porę do cytoplazmy. Mówiąc prościej, TREX-2 działa jak inteligentna stacja dokująca, która zarówno rozpoznaje nadchodzący ładunek, jak i uruchamia mechanizm napędzany paliwem, by odczepić go od eskorty i przekazać nosicielowi wychodzącemu.
Dlaczego ta mikroskopijna choreografia ma znaczenie
Praca ta wyjaśnia, na poziomie niemal atomowym, jak komórki ludzkie używają zachowanego przełącznika molekularnego do kontrolowania, które komunikaty mRNA opuszczają jądro. UAP56 wyłania się jako centralny kontroler ruchu, którego stan jest odczytywany i resetowany przez TREX-2, zapewniając, że tylko prawidłowo zapakowane mRNA trafiają do aparatu eksportowego. Ponieważ błędy w tej ścieżce wiążą się z niestabilnością genomu i chorobami, zrozumienie tej choreografii daje podstawę do badania, jak mutacje w tych czynnikach mogą powodować schorzenia i, w dłuższej perspektywie, do projektowania sposobów selektywnego regulowania eksportu mRNA w terapiach.
Cytowanie: Gong, X., Tao, R., Ge, X. et al. Molecular insights into mRNA export regulation by the human TREX-2 complex. Nat Commun 17, 3244 (2026). https://doi.org/10.1038/s41467-026-70088-w
Słowa kluczowe: eksport mRNA, kompleks TREX-2, helikaza UAP56, otwór jądrowy, przetwarzanie RNA