Clear Sky Science · nl
Molculaire inzichten in de regulatie van mRNA-export door het menselijke TREX-2-complex
Hoe cellen beslissen welke boodschappen de kern mogen verlaten
Elke seconde sturen onze cellen duizenden genetische “boodschappen” in de vorm van boodschapper-RNA (mRNA) uit die aangeven welke eiwitten geproduceerd moeten worden. Onjuiste boodschappen doorlaten — of ze op het verkeerde moment vrijgeven — kan ernstige gevolgen hebben, van ontwikkelingsstoornissen tot kanker. Deze studie onthult hoe een belangrijk poortwachterscomplex bij het nucleaire pore, genaamd TREX-2, samenwerkt met een moleculaire motor, UAP56, om te bepalen wanneer een mRNA-pakket klaar is om de kern te verlaten.
Van losse tekst tot verzegelde boodschap
Voordat een mRNA de kern kan verlaten, moet het zorgvuldig verwerkt en omhuld worden met vele eiwitten om een stabiel, exportklaar deeltje te vormen. Eerder werk toonde dat één assemblagelijn, bekend als TREX, het mRNA verpakt en het klemzet samen met UAP56 — een motorachtig eiwit dat cellulaire brandstof (ATP) gebruikt en sterk aan RNA bindt. Een tweede complex, TREX-2, zit aangemeerd bij nucleaire pores — de doorgangen in het nucleaire membraan — en is essentieel voor export. Hoe TREX-2 het UAP56-gebonden mRNA herkent en hoe de boodschap wordt doorgegeven aan de uiteindelijke exportdrager, bleef echter onduidelijk. 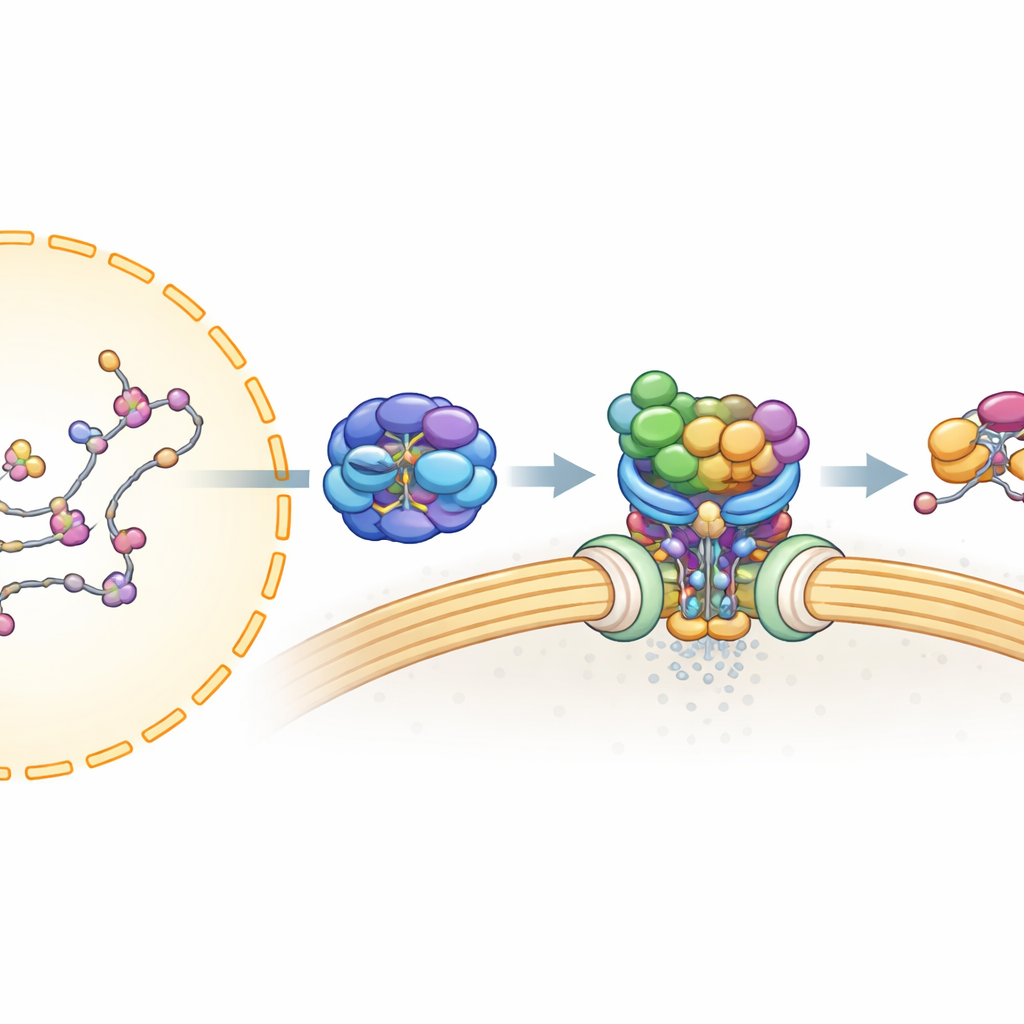
Een moleculaire handdruk bij de nucleaire poort
De auteurs gebruikten menselijke cellen, eiwitzuivering en massaspectrometrie om in kaart te brengen welke eiwitten fysiek met TREX-2 associëren. Ze vonden dat UAP56, maar niet de meeste andere TREX-componenten, sterk bindt aan het kernskelet van TREX-2, opgebouwd uit eiwitten genaamd GANP, PCID2 en DSS1. Dit wees op UAP56 als een toegewijde adaptor die verpakt mRNA naar TREX-2 brengt. vervolgexperimenten toonden aan dat UAP56 en TREX-2 alleen een strakke complexvorming aangaan wanneer UAP56 een nucleotide vasthoudt (een brandstofgerelateerd molecuul zoals ATP of ADP), wat suggereert dat TREX-2 een specifieke “geladen” toestand van UAP56 in de exportcyclus herkent.
De poortwachter op atomair niveau zien
Om deze interactie op hoge resolutie te begrijpen, ging het team over op cryo-elektronenmicroscopie, die ingevroren moleculen afbeeldt en onderzoekers in staat stelt hun driedimensionale vormen te reconstrueren. Ze bepaalden structuren van TREX-2 alleen en gebonden aan UAP56. De kern van TREX-2 vormt een V-vormige cradle, waarbij GANP en PCID2 de armen vormen en DSS1 zich langs één zijde nestelt. UAP56 zit in deze cradle als een bal in een sling, en maakt voornamelijk contact met TREX-2 via twee flexibele lusjes aan zijn uiterste begin (de N-terminale regio). Deze lusjes reiken in een positief geladen groef op GANP en PCID2 en vergrendelen UAP56 op het V-vormige oppervlak. Mutaties van sleutelaminozuren in deze lusjes — in feite het afknippen of botmaken van de haakpuntjes — verzwakten of sloegen de binding grotendeels af, wat bevestigde dat dit kleine gebied van UAP56 de belangrijkste dockingsite voor TREX-2 is.
Hoe TREX-2 de motor dwingt los te laten
Verrassend genoeg zit UAP56, wanneer het aan TREX-2 gebonden is, vast in een “open” houding die RNA niet vasthoudt, ook al was het complex met RNA aanwezig samengesteld. De structuur verklaart waarom: een korte lus van GANP, het activeringslusje genoemd, klemt zich tussen de twee lobben van UAP56 die normaal gesproken de RNA-streng klemmen. Deze lus reikt ook in de ATP-bindingspocket van UAP56 en duwt sleutelzijgroepen die normaal ATP waarnemen. Biochemische assays toonden aan dat TREX-2 de ATPase-activiteit van UAP56 — het tempo waarin het ATP verbrandt — met ongeveer tienvoud vergroot, maar alleen wanneer het activeringslusje en de N-terminale dockingslusjes intact zijn. In gelelektroforese gebaseerde RNA-loslaatexperimenten scheurde TREX-2 actief RNA van UAP56, terwijl een mutant TREX-2 met één gewijzigd residu in het activeringslusje een veel zwakker effect had, en een UAP56-mutant die zijn dockingsegment miste helemaal niet reageerde. 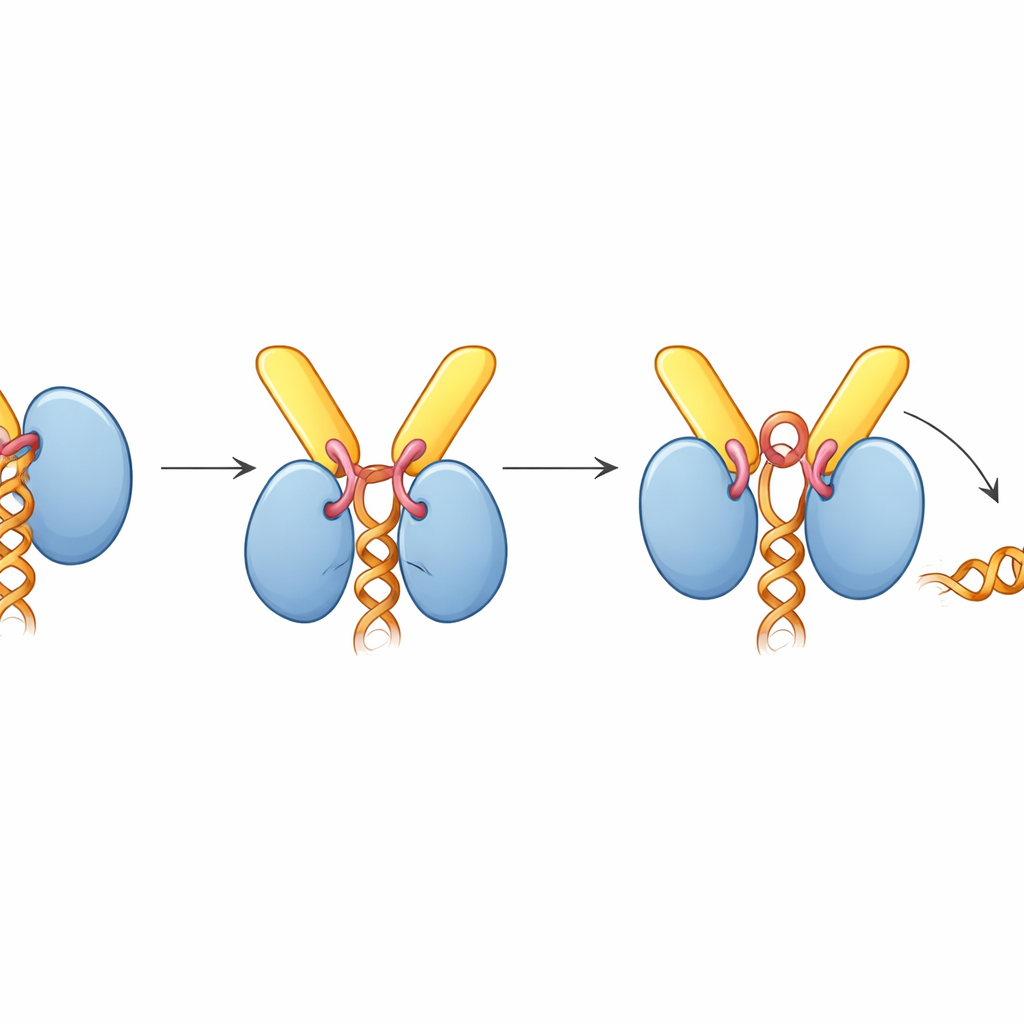
Een gecoördineerde overdracht van genetische vracht
Op basis van deze bevindingen stellen de auteurs een verenigd model voor mRNA-export voor. TREX en UAP56 verpakken en klemmen eerst mRNA in compacte deeltjes binnen de kern. Deze deeltjes diffunderen vervolgens totdat het UAP56-“handvat” wordt gevangen door TREX-2 bij het nucleaire pore. Het activeringslusje van TREX-2 duwt UAP56 in een conformatie die ATP verbruikt en zijn RNA-greep opendoet, waardoor het mRNA vrijkomt. De vrijgekomen boodschap wordt onmiddellijk opgepikt door de uiteindelijke exportreceptor (NXF1–NXT1), die eveneens door TREX-2 wordt gerekruteerd, en vervolgens door de poort naar het cytoplasma vervoerd. In eenvoudige termen fungeert TREX-2 als een slimme aanlegplaats die zowel binnenkomende lading herkent als een brandstofgestuurde mechaniek activeert om die van zijn escort los te maken en aan de uitgaande drager over te geven.
Waarom deze microscopische choreografie ertoe doet
Dit werk verklaart, tot op bijna atomair niveau, hoe menselijke cellen een geconserveerde moleculaire schakelaar gebruiken om te controleren welke mRNA-boodschappen de kern ontsnappen. UAP56 komt naar voren als een centrale verkeersregelaar waarvan de toestand wordt afgelezen en gereset door TREX-2, wat ervoor zorgt dat alleen correct verpakt mRNA aan de exportmachinerie wordt overgedragen. Omdat fouten in dit pad verband houden met genome-instabiliteit en ziekte, biedt begrip van deze choreografie een basis om te onderzoeken hoe mutaties in deze factoren ziekte kunnen veroorzaken en, op de lange termijn, om manieren te ontwerpen om mRNA-export selectief te reguleren in therapieën.
Bronvermelding: Gong, X., Tao, R., Ge, X. et al. Molecular insights into mRNA export regulation by the human TREX-2 complex. Nat Commun 17, 3244 (2026). https://doi.org/10.1038/s41467-026-70088-w
Trefwoorden: mRNA-export, TREX-2-complex, UAP56-helicase, nucleair pore, RNA-verwerking