Clear Sky Science · en
Molecular insights into mRNA export regulation by the human TREX-2 complex
How cells decide which messages may leave the nucleus
Every second, our cells send out thousands of genetic “messages” in the form of messenger RNA (mRNA) that tell the rest of the cell which proteins to make. Letting the wrong messages through—or releasing them at the wrong time—can lead to serious problems, from developmental defects to cancer. This study uncovers how a key gatekeeping complex at the nuclear pore, called TREX-2, works together with a molecular motor protein, UAP56, to decide when an mRNA package is ready to leave the nucleus.
From loose script to sealed message
Before an mRNA can exit the nucleus, it must be carefully processed and wrapped with many proteins to form a stable, export-ready particle. Earlier work showed that one assembly line, known as TREX, packages the mRNA and clamps it together with UAP56, a motor-like protein that uses cellular fuel (ATP) and binds tightly to RNA. A second complex, TREX-2, sits docked at nuclear pores—the gateways in the nuclear membrane—and is known to be essential for export. But how TREX-2 recognizes the UAP56-bound mRNA, and how the message is passed on to the final export carrier, had remained unclear. 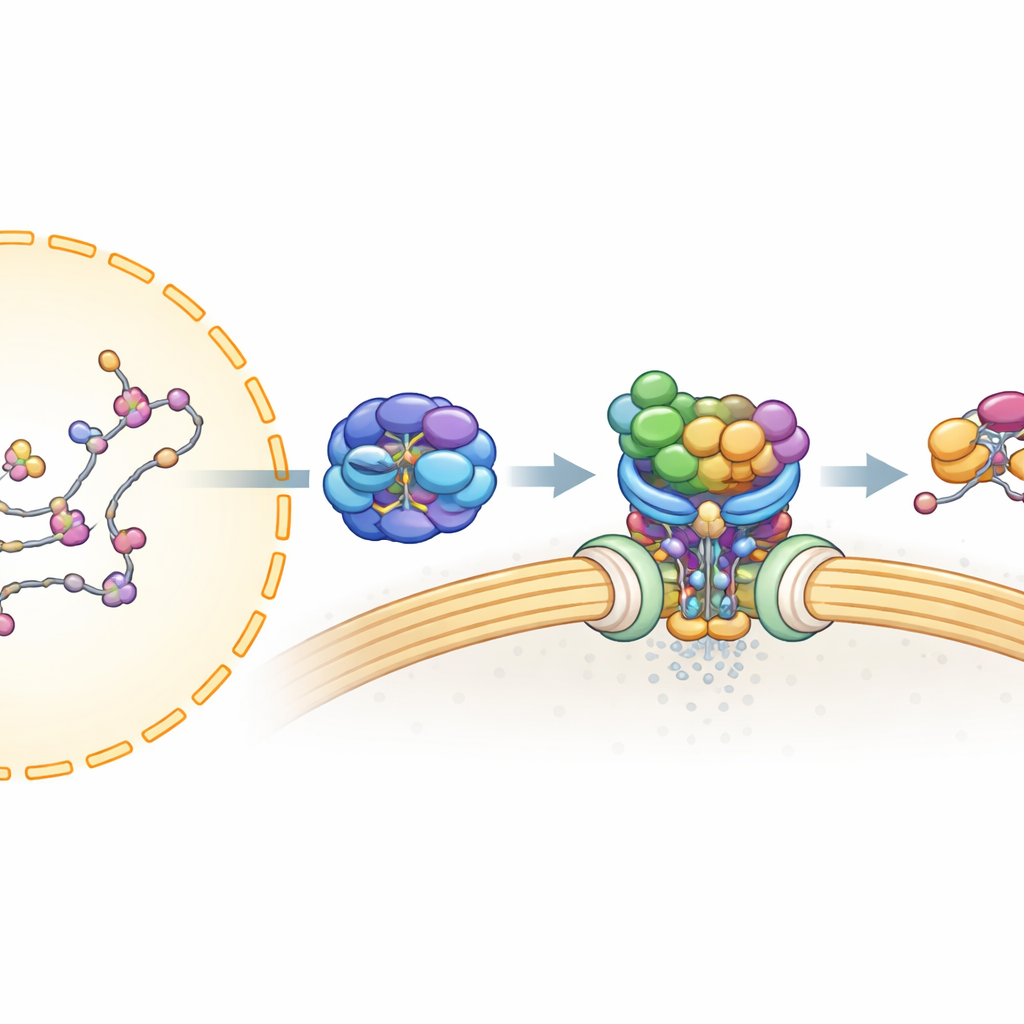
A molecular handshake at the nuclear gate
The authors used human cells, protein purification, and mass spectrometry to map which proteins physically associate with TREX-2. They found that UAP56, but not most other TREX components, strongly binds to TREX-2’s core scaffold, built from proteins called GANP, PCID2, and DSS1. This pointed to UAP56 as a dedicated adaptor that brings packaged mRNA to TREX-2. Follow-up experiments showed that UAP56 and TREX-2 only form a tight complex when UAP56 holds a nucleotide (a fuel-related molecule such as ATP or ADP), suggesting that TREX-2 recognizes a specific “loaded” state of UAP56 during the export cycle.
Seeing the gatekeeper in atomic detail
To understand this interaction at high resolution, the team turned to cryo–electron microscopy, which images flash-frozen molecules and lets researchers reconstruct their 3D shapes. They solved structures of TREX-2 alone and bound to UAP56. TREX-2’s core forms a V-shaped cradle, with GANP and PCID2 making the arms and DSS1 nestling along one side. UAP56 sits in this cradle like a ball in a sling, contacting TREX-2 mainly through two flexible loops at its extreme beginning (its N-terminal region). These loops reach into a positively charged groove on GANP and PCID2, locking UAP56 onto the V-shaped surface. Mutating key amino acids in these loops—in effect snipping or dulling the tips of the hooks—greatly weakened or abolished binding, confirming that this small region of UAP56 is the main docking site for TREX-2.
How TREX-2 forces the motor to let go
Surprisingly, when TREX-2 is bound, UAP56 is caught in an “open” pose that does not hold RNA, even though the complex was assembled with RNA present. The structure reveals why: a short loop from GANP, dubbed the activating loop, wedges itself between the two lobes of UAP56 that normally clamp the RNA strand. This loop also reaches into UAP56’s ATP-binding pocket, nudging key side chains that normally sense ATP. Biochemical assays showed that TREX-2 greatly boosts UAP56’s ATPase activity—the rate at which it burns ATP—by about tenfold, but only when the activating loop and the N-terminal docking loops are intact. In gel-based RNA release experiments, TREX-2 actively stripped RNA off UAP56, whereas a mutant TREX-2 with a single altered residue in the activating loop had a much weaker effect, and a UAP56 mutant lacking its docking segment did not respond at all. 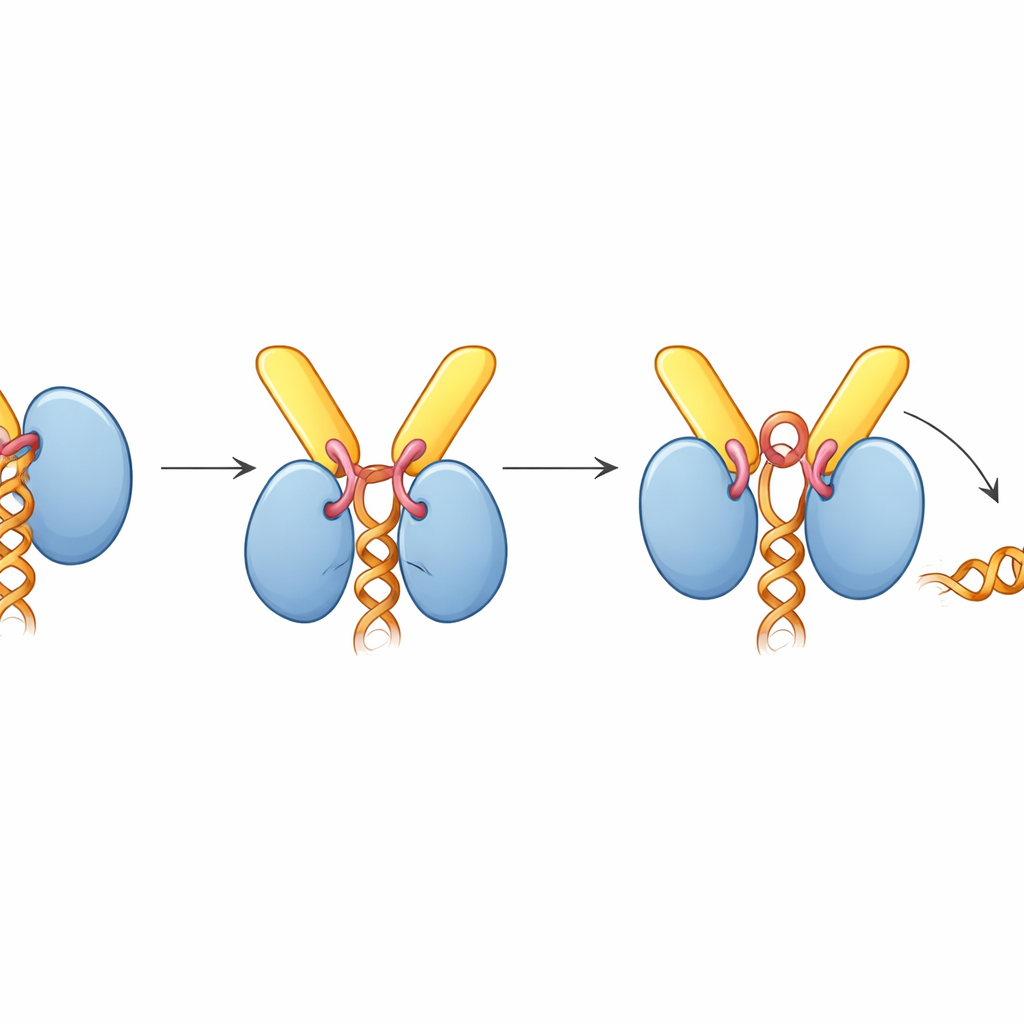
A coordinated handoff of genetic cargo
Putting these findings together, the authors propose a unified model for mRNA export. TREX and UAP56 first package and clamp mRNA into compact particles inside the nucleus. These particles then diffuse until the UAP56 “handle” is captured by TREX-2 at the nuclear pore. TREX-2’s activating loop pushes UAP56 into a conformation that burns ATP and pries open its RNA grip, releasing the mRNA. The freed message is immediately caught by the final export receptor (NXF1–NXT1), which is also recruited by TREX-2, and then ferried through the pore to the cytoplasm. In simple terms, TREX-2 acts as a smart docking station that both recognizes incoming cargo and triggers a fuel-driven mechanism to unclip it from its escort and pass it to the outbound carrier.
Why this microscopic choreography matters
This work explains, at near-atomic detail, how human cells use a conserved molecular switch to control which mRNA messages escape the nucleus. UAP56 emerges as a central traffic controller whose state is read and reset by TREX-2, ensuring that only properly packaged mRNA is handed to the export machinery. Because errors in this pathway are linked to genome instability and disease, understanding this choreography provides a foundation for exploring how mutations in these factors might cause illness and, in the long term, for designing ways to selectively tune mRNA export in therapies.
Citation: Gong, X., Tao, R., Ge, X. et al. Molecular insights into mRNA export regulation by the human TREX-2 complex. Nat Commun 17, 3244 (2026). https://doi.org/10.1038/s41467-026-70088-w
Keywords: mRNA export, TREX-2 complex, UAP56 helicase, nuclear pore, RNA processing