Clear Sky Science · sv
Molekylära insikter i regleringen av mRNA-export genom det humana TREX-2-komplexet
Hur celler avgör vilka budskap som får lämna kärnan
Varje sekund skickar våra celler ut tusentals genetiska ”budskap” i form av budbärar-RNA (mRNA) som talar om för resten av cellen vilka proteiner som ska tillverkas. Att släppa igenom fel budskap — eller frigöra dem vid fel tidpunkt — kan leda till allvarliga problem, från utvecklingsstörningar till cancer. Denna studie avslöjar hur ett centralt grindvaktskomplex vid kärnporen, kallat TREX-2, samarbetar med ett molekylärt motorprotein, UAP56, för att avgöra när ett mRNA-paket är redo att lämna kärnan.
Från lös manuskript till förseglat meddelande
Innan ett mRNA kan lämna kärnan måste det noggrant bearbetas och omslutas av många proteiner för att bilda en stabil, exportredo partikel. Tidigare arbete visade att en sådan monteringslinje, känd som TREX, paketerar mRNA och klämmer ihop det med UAP56, ett motorliknande protein som använder cellulärt bränsle (ATP) och binder tätt till RNA. Ett andra komplex, TREX-2, sitter förankrat vid kärnporerna — portarna i kärnmembranet — och är känt för att vara nödvändigt för export. Men hur TREX-2 känner igen det UAP56-bundna mRNA:t, och hur budskapet överlämnas till den slutliga exportbäraren, hade hittills varit oklart. 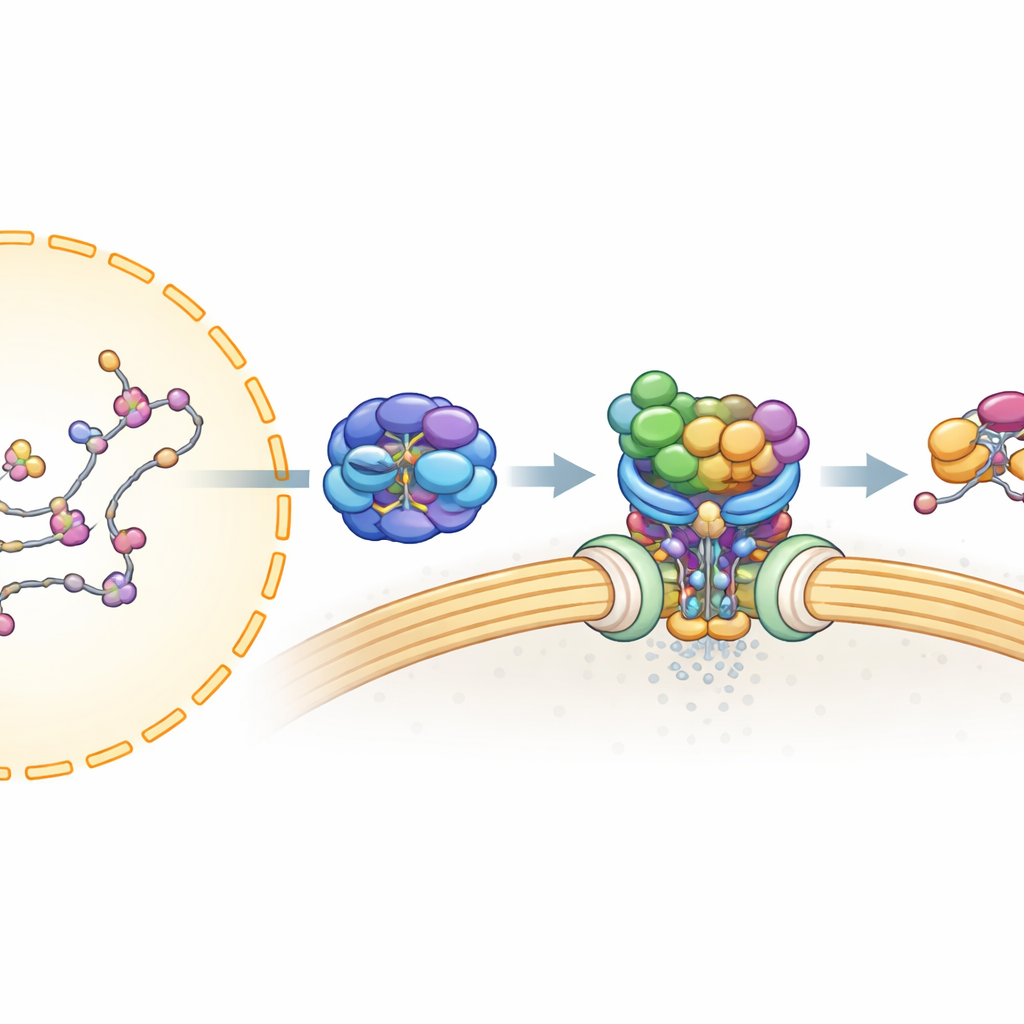
En molekylär handskakning vid kärnporten
Författarna använde humana celler, proteinrening och masspektrometri för att kartlägga vilka proteiner som fysiskt associerar med TREX-2. De fann att UAP56, men inte de flesta andra TREX-komponenter, binder starkt till TREX-2:s kärnskellett, uppbyggt av proteinerna GANP, PCID2 och DSS1. Detta pekade på UAP56 som en dedikerad adapter som förde det paketerade mRNA:t till TREX-2. Uppföljande experiment visade att UAP56 och TREX-2 endast bildar ett tätt komplex när UAP56 håller en nukleotid (en bränslerelaterad molekyl såsom ATP eller ADP), vilket tyder på att TREX-2 känner igen ett specifikt ”laddat” tillstånd av UAP56 under exportcykeln.
Att se grindvakten i atomär detalj
För att förstå denna interaktion med hög upplösning använde teamet kryoelektronmikroskopi, som avbildar blixtfrusna molekyler och låter forskare rekonstruera deras 3D-former. De löste strukturer av TREX-2 ensam och bundet till UAP56. TREX-2:s kärna bildar en V-formad vagga, med GANP och PCID2 som armar och DSS1 vilande längs ena sidan. UAP56 sitter i denna vagga som en boll i en slunga, och kontaktar TREX-2 främst genom två flexibla slingor vid dess yttersta början (dess N-terminala region). Dessa slingor når in i en positivt laddad fåra på GANP och PCID2 och låser UAP56 på den V-formade ytan. Mutationer i viktiga aminosyror i dessa slingor — i praktiken att klippa eller stumpa kroktopparna — försvagade eller upphävde bindningen avsevärt, vilket bekräftade att detta lilla område av UAP56 är huvudankringsplatsen för TREX-2.
Hur TREX-2 tvingar motorn att släppa taget
Överraskande nog fångas UAP56 i en ”öppen” ställning när TREX-2 är bundet, vilket inte håller RNA, trots att komplexet monterades med RNA närvarande. Strukturen visar varför: en kort slinga från GANP, som kallats aktiveringsslingan, kilas in mellan UAP56:s två lobar som normalt klämmer runt RNA-strängen. Denna slinga når också in i UAP56:s ATP-bindningsficka och petar på viktiga sidokedjor som normalt känner av ATP. Biokemiska tester visade att TREX-2 kraftigt ökar UAP56:s ATPasaktivitet — takten vid vilken det förbrukar ATP — med ungefär tiofald, men endast när aktiveringsslingan och de N-terminala dockningsslingorna är intakta. I gelbaserade experiment för RNA-frisättning strippade TREX-2 aktivt RNA från UAP56, medan en mutant TREX-2 med en enda ändrad rest i aktiveringsslingan hade en mycket svagare effekt, och en UAP56-mutant utan sin dockningssektion svarade inte alls. 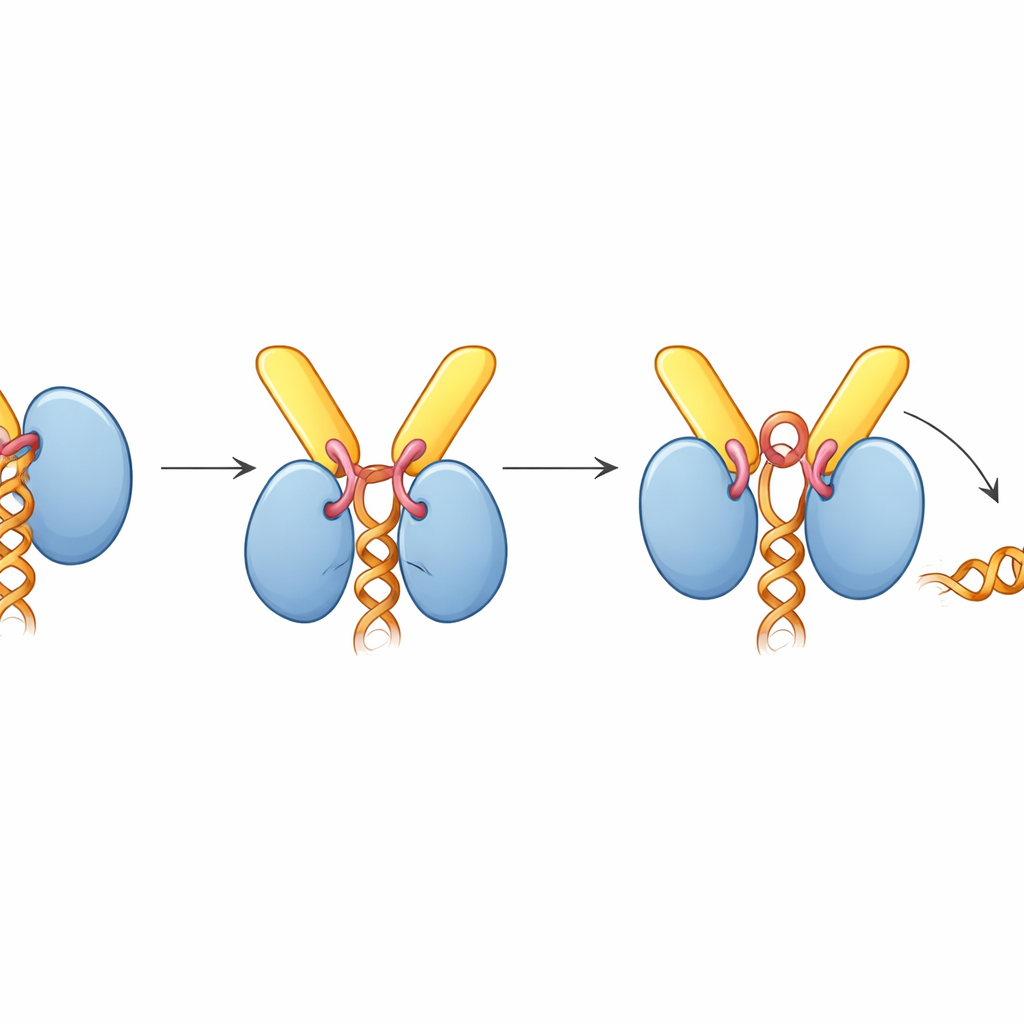
En samordnad överlämning av genetiskt last
Sammantaget föreslår författarna en enhetlig modell för mRNA-export. TREX och UAP56 paketerar och klämmer först mRNA till kompakta partiklar inne i kärnan. Dessa partiklar diffunderar sedan tills UAP56-”handtaget” fångas av TREX-2 vid kärnporen. TREX-2:s aktiveringsslinga skjuter UAP56 i en konformation som förbrukar ATP och sliter upp dess RNA-grepp, vilket frigör mRNA:t. Det frigjorda budskapet fångas omedelbart av den slutliga exportreceptorn (NXF1–NXT1), som också rekryteras av TREX-2, och förs sedan genom poren till cytoplasman. Enkelt uttryckt fungerar TREX-2 som en smart dokningsstation som både känner igen inkommande last och triggar en bränsledriven mekanism för att lossna den från dess eskort och föra över den till den avgående bäraren.
Varför denna mikroskopiska koreografi är viktig
Detta arbete förklarar, på nästan atomär nivå, hur humana celler använder en bevarad molekylär brytare för att kontrollera vilka mRNA-budskap som undkommer kärnan. UAP56 framträder som en central trafikreglerare vars tillstånd avläses och återställs av TREX-2, vilket säkerställer att endast korrekt paketerat mRNA överlämnas till exportmaskineriet. Eftersom fel i denna bana kopplats till genominstabilitet och sjukdom ger förståelsen av denna koreografi en grund för att utforska hur mutationer i dessa faktorer kan orsaka sjukdom, och på längre sikt för att utforma sätt att selektivt påverka mRNA-export i terapier.
Citering: Gong, X., Tao, R., Ge, X. et al. Molecular insights into mRNA export regulation by the human TREX-2 complex. Nat Commun 17, 3244 (2026). https://doi.org/10.1038/s41467-026-70088-w
Nyckelord: mRNA-export, TREX-2-komplex, UAP56-helikas, kärnpore, RNA-processorering