Clear Sky Science · ru
Молекулярные механизмы регуляции экспорта мРНК комплексом TREX-2 у человека
Как клетки решают, какие сообщения могут покинуть ядро
Каждую секунду наши клетки отправляют тысячи генетических «сообщений» в виде матричной РНК (мРНК), которые указывают остальной части клетки, какие белки синтезировать. Пропуск неверных сообщений — или выпуск их в неподходящее время — может привести к серьёзным последствиям, от нарушений развития до рака. В этом исследовании раскрывается, как ключевой «шлюзовой» комплекс у ядерной поры, называемый TREX-2, действует совместно с молекулярным мотором UAP56, чтобы определить, когда упаковка мРНК готова покинуть ядро.
От разрозненного сценария к запечатанному сообщению
Прежде чем мРНК сможет выйти из ядра, её нужно тщательно обработать и окружить множеством белков, чтобы получить стабильную частицу, готовую к экспорту. Ранее показали, что одна «сборочная линия», известная как TREX, упаковывает мРНК и фиксирует её с помощью UAP56 — мотороподобного белка, который использует клеточное топливо (АТФ) и прочно связывается с РНК. Второй комплекс, TREX-2, расположен у ядерных пор — ворот в ядерной оболочке — и необходим для экспорта. Но оставалось неясным, как TREX-2 распознаёт мРНК, связанную с UAP56, и как сообщение передаётся окончательному «перевозчику» для экспорта. 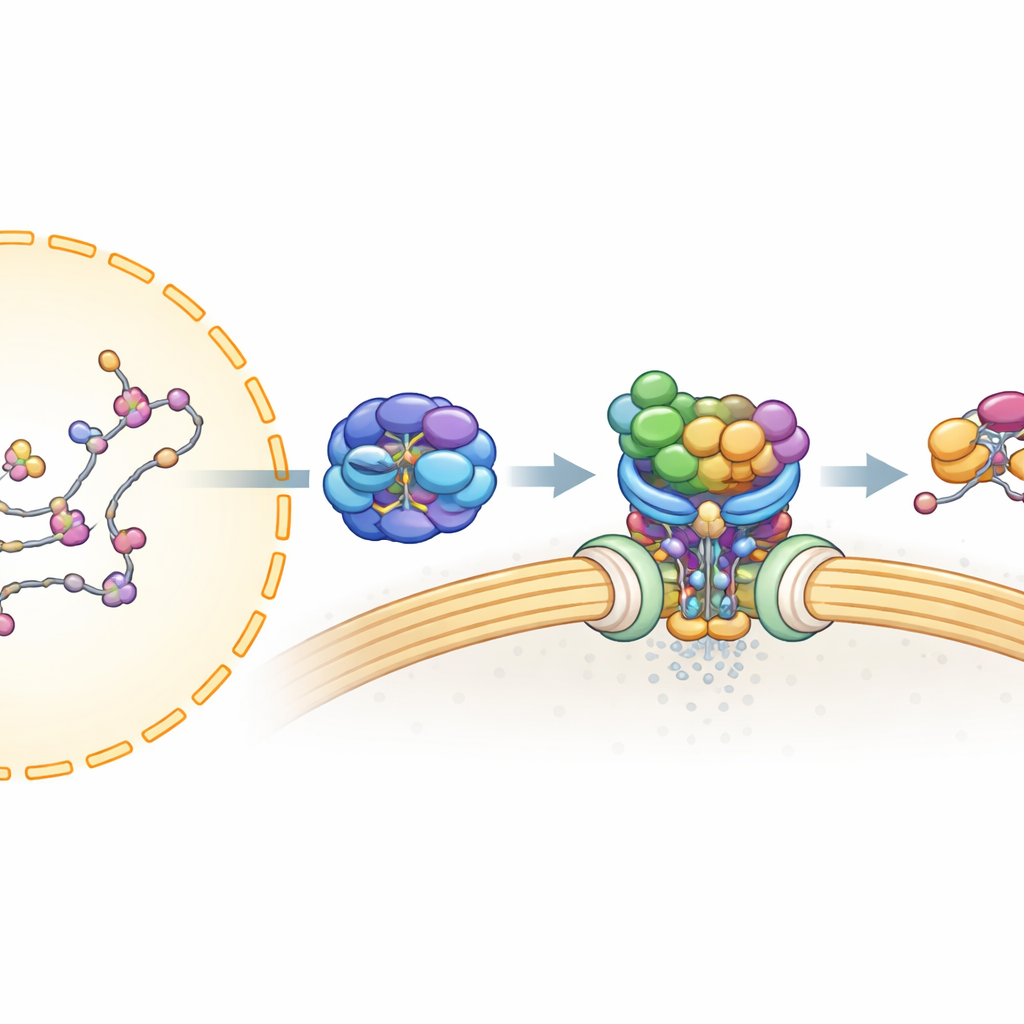
Молекулярное рукопожатие у ядерных ворот
Авторы использовали клетки человека, очистку белков и масс-спектрометрию, чтобы отобразить, какие белки физически ассоциируются с TREX-2. Они обнаружили, что UAP56, в отличие от большинства других компонентов TREX, сильно связывается с основным каркасом TREX-2, сформированным белками GANP, PCID2 и DSS1. Это указывало на UAP56 как на специализированный адаптер, который приносит упакованную мРНК к TREX-2. Последующие эксперименты показали, что UAP56 и TREX-2 образуют прочный комплекс только тогда, когда UAP56 удерживает нуклеотид (молекулу, связанную с топливом, такую как АТФ или АДФ), что предполагает, что TREX-2 распознаёт специфическое «заряженное» состояние UAP56 в ходе цикла экспорта.
Взгляд на стража в атомном разрешении
Чтобы понять это взаимодействие с высокой точностью, команда применила криоэлектронную микроскопию, которая позволяет получать изображения быстро замороженных молекул и реконструировать их трёхмерные формы. Они получили структуры TREX-2 в свободном виде и в комплексе с UAP56. Ядро TREX-2 формирует V-образную «люльку»: GANP и PCID2 образуют «руки», а DSS1 располагается вдоль одной стороны. UAP56 сидит в этой люльке как мяч в петле, контактируя с TREX-2 главным образом через два гибких петлевых участка в его крайнем N-конце. Эти петли входят в положительно заряженную борозду на GANP и PCID2, фиксируя UAP56 на V-образной поверхности. Мутации ключевых аминокислот в этих петлях — по сути укорачивание или притупление «крючков» — значительно ослабляли или устраняли связывание, что подтверждает, что именно этот небольшой участок UAP56 служит основным сайтом докинга для TREX-2.
Как TREX-2 заставляет мотор отпустить РНК
Удивительно, но при связывании с TREX-2 UAP56 оказывается в «открытой» конформации, не удерживающей РНК, несмотря на то, что комплекс собирали в присутствии РНК. Структура объясняет это: короткая петля от GANP, названная активирующей петлёй, вклинивается между двумя долями UAP56, которые обычно захватывают РНК. Эта петля также проникает в АТФ-связывающий карман UAP56, смещая боковые цепи аминокислот, которые обычно «чувствуют» АТФ. Биохимические анализы показали, что TREX-2 значительно усиливает АТФазную активность UAP56 — скорость расходования АТФ — примерно в десять раз, но только при неповреждённой активирующей петле и N-концевых доковых петлях. В гель-основных экспериментах по освобождению РНК TREX-2 активно снимал РНК с UAP56, тогда как мутант TREX-2 с изменённой одной аминокислотой в активирующей петле имел значительно более слабый эффект, а мутант UAP56, лишённый докового сегмента, вообще не реагировал. 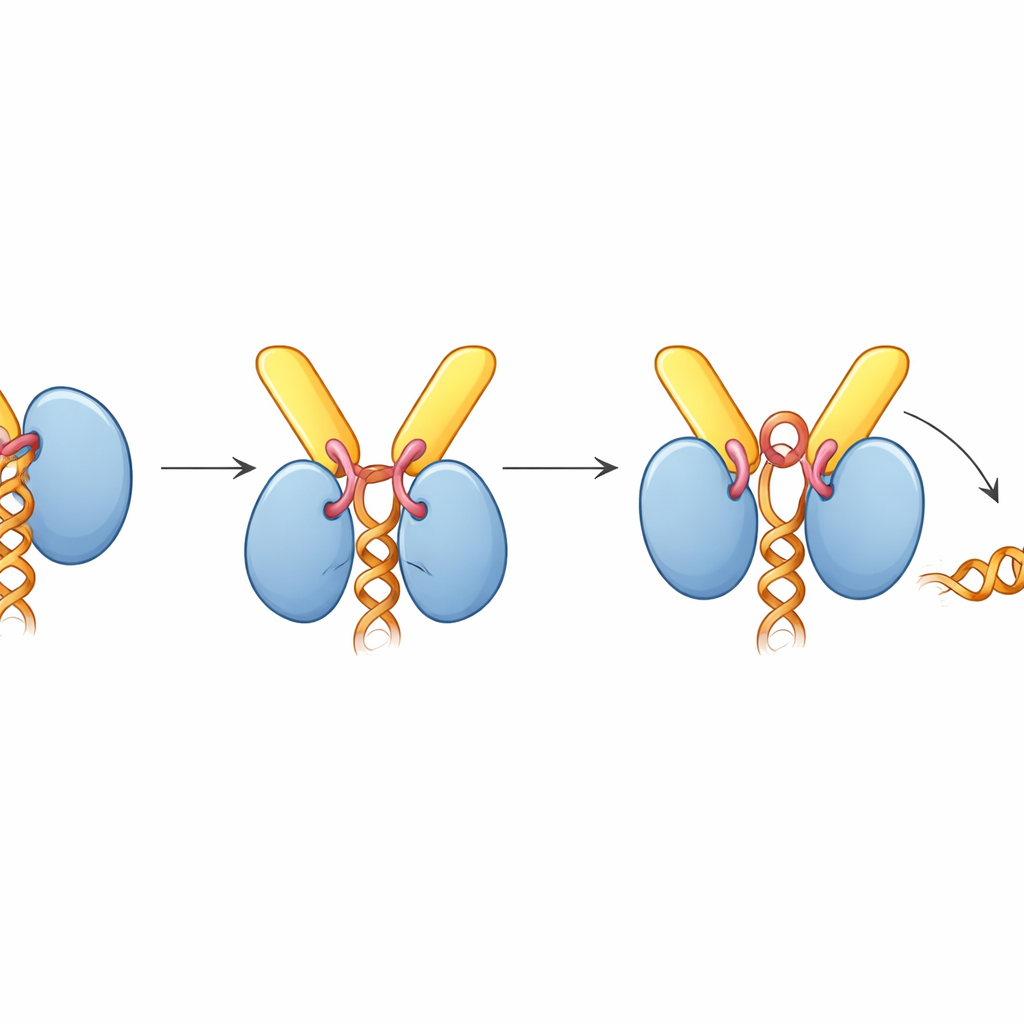
Скоординированная передача генетического груза
Объединив эти результаты, авторы предлагают единый механизм экспорта мРНК. TREX и UAP56 сначала упаковывают и фиксируют мРНК в компактные частицы внутри ядра. Эти частицы затем диффундируют до тех пор, пока «ручка» UAP56 не будет захвачена TREX-2 у ядерной поры. Активирующая петля TREX-2 переводит UAP56 в конформацию, при которой тот расходует АТФ и раскрывает захват РНК, освобождая мРНК. Освобождённое сообщение тут же перехватывается окончательным рецептором экспорта (NXF1–NXT1), который также привлекается TREX-2, и затем транспортируется через пору в цитоплазму. Проще говоря, TREX-2 действует как интеллектуальная док-станция, которая и распознаёт приходящий груз, и запускает топливозависимый механизм, чтобы отщёлкнуть его от эскорта и передать наружному переносчику.
Почему этот микроскопический танец важен
Эта работа объясняет с почти атомной детализацией, как клетки человека используют консервативный молекулярный переключатель для контроля того, какие сообщения мРНК покидают ядро. UAP56 предстает в роли центрального диспетчера трафика, состояние которого считывается и сбрасывается TREX-2, что гарантирует передачу на экспорт только правильно упакованной мРНК. Поскольку ошибки в этом пути связаны с нестабильностью генома и заболеваниями, понимание этой «хореографии» даёт основу для изучения того, как мутации в этих факторах могут вызывать болезни и, в долгосрочной перспективе, для разработки способов селективной модуляции экспорта мРНК в терапевтических целях.
Цитирование: Gong, X., Tao, R., Ge, X. et al. Molecular insights into mRNA export regulation by the human TREX-2 complex. Nat Commun 17, 3244 (2026). https://doi.org/10.1038/s41467-026-70088-w
Ключевые слова: экспорт мРНК, комплекс TREX-2, хеликаза UAP56, ядерная пора, процессинг РНК