Clear Sky Science · sv
Icke-enzymatisk felkorrigering i självreplikerare utan extern energiförsörjning
Varför det spelar roll att kopiera livets kod utan enzymer
Innan cellerna utvecklade sina sofistikerade molekylära maskiner måste tidigt genetiskt material på jorden kopiera sig självt enbart med enkel kemi. Att kopiera långa kedjor av "bokstäver"—som i DNA och RNA—utan att göra för många misstag är dock extremt svårt. Denna artikel undersöker hur relativt enkla molekyler skulle kunna kopiera sig själva korrekt, utan enzymer och utan extra bränsletillförsel som ATP, och erbjuder en konkret väg för hur livets första genetiska system kunde uppstå och bestå.
En gåta om misstag och livets uppkomst
Moderna celler använder specialiserade proteiner för att korrekturläsa DNA och åtgärda fel, och förbrukar kemisk energi för detta. Dessa enzymer är emellertid alltför komplexa för att ha funnits på den tidiga jorden. Utan dem skulle primitiva självkopierande strängar ha ackumulerat fel så snabbt att användbar information inte kunde bevaras från en generation till nästa. Befintliga teorier antar antingen samhällen av ömsesidigt hjälpsamma molekyler eller förlitar sig på finjusterade miljöer eller extra energikällor. Författarna frågar istället: skulle en enda självreplikerande kedja kunna korrigera sina egna kopieringsfel med enbart den energi som redan driver dess tillväxt?
En ensidig skjuts som styr tillväxten
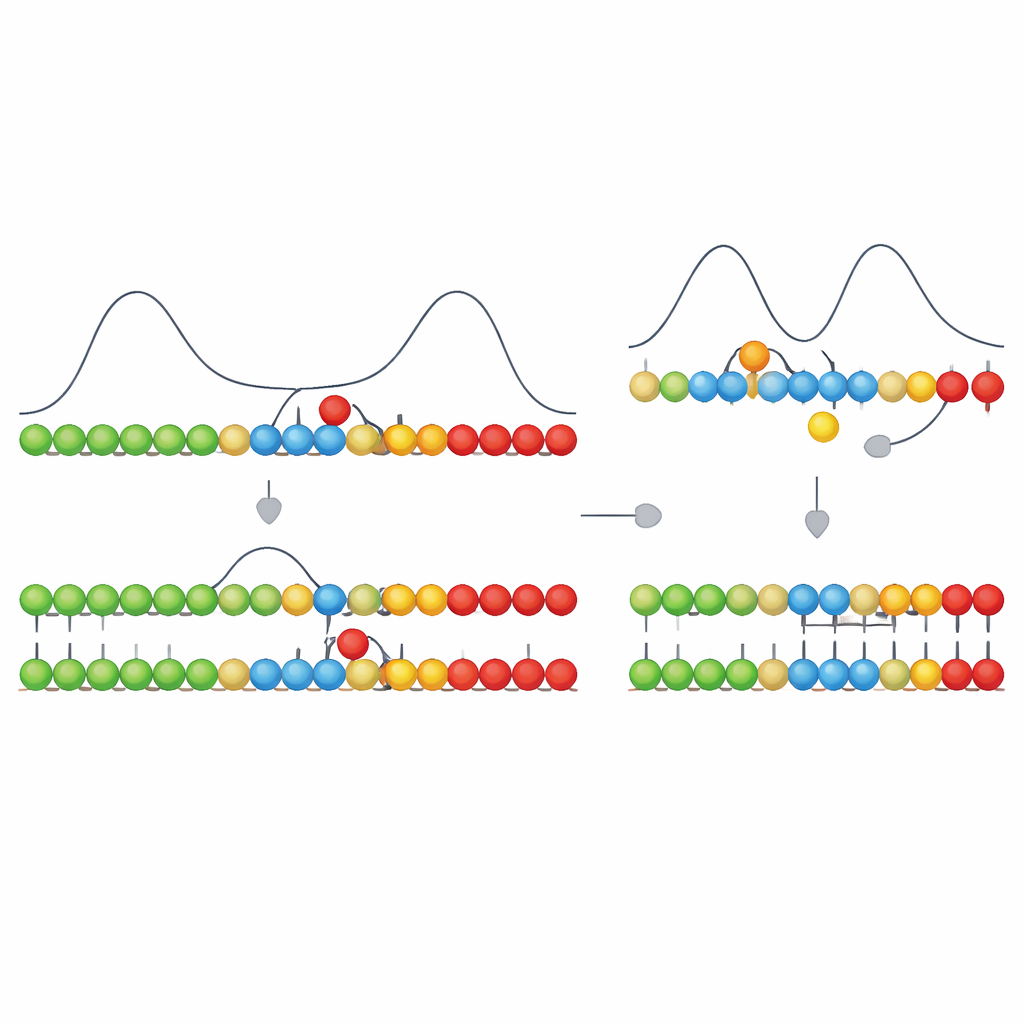
Författarna bygger vidare på en kinetisk egenskap de kallar asymmetrisk kooperativitet. Föreställ dig en mallsträng där nya byggstenar kortvarigt fastnar vid svaga bindningar. När rätt typ av byggsten binder på en position gör det det lättare för en annan byggsten att binda på en viss sida (till exempel åt höger) och svårare för den befintliga bindningen att falla isär. På motsatt sida gör det saker något svårare, vilket i praktiken driver tillväxten i en enda riktning längs mallen. I deras modell har korrekta parningar denna riktade effekt, medan felaktiga inte har det. Denna enkla regel får korrekt parade segment att växa snabbt i en riktning, medan felställen bromsar tillväxten vid sin plats och destabiliserar närliggande bindningar.
Att omvandla tidsdifferenser till färre fel
På egen hand skulle denna riktade bromsning bara skapa tillfälliga skillnader: korrekta segment fortskrider mjukt; segment med en felaktig byggsten pausar och tenderar att veckas upp. Nyckelsteget är att intilliggande enheter på den växande strängen också kan bilda starka, nästan irreversibla kemiska länkar med varandra. Att bilda dessa länkar är energetiskt mycket fördelaktigt, men kan vara snabbt eller långsamt beroende på kemin. Författarna visar att när detta länkbildande steg går tillräckligt snabbt—i samma tidskala som den korta fördröjning som orsakas av en mismatch—kommer det att föredra att "låsa in" sträckor som råkade vara korrekta, innan en felaktig enhet hinner stabiliseras. Om länkbildningen är för långsam har allt återutjämnats innan en länk bildas, och systemet kan inte längre skilja rätt från fel.
Att simulera en primitiv kopieringsmaskin
För att utforska idén behandlar gruppen varje kort mall som en kedja av positioner som kan vara obundna, korrekt parade eller felaktigt parade. De använder sedan en Markovkjedjemodell—ett standardmatematiskt verktyg för slumpmässiga steg-för-steg-processer—för att följa alla möjliga vägar när baspar bildas och bryts. Genom att variera den riktade biasen, hastigheterna för parning och upplösning samt hastigheten för kovalent länkning beräknar de både hur ofta en slutlig kopierad sträng innehåller ett fel och hur lång tid kopiering tar. De finner att stark envägskooperativitet och tillräckligt snabb länkbildning tillsammans kan pressa ner felhastigheten från en rent termodynamisk baslinje på ungefär ett fel på hundra till omkring ett på tio tusen, jämförbart med vad som ses hos verkliga DNA-polymeraser under deras "passiva" basvalsstadium, innan extra korrekturläsning träder in.
Mönster som liknar verklig biologi
Påfallande nog återger denna avskalade modell flera egenskaper som observeras vid modern DNA-kopiering. När en mismatch uppträder saktar den simulerade strängtillväxten dramatiskt—en form av "stallning" som också ses i experiment. Mismatcher leder till en ökad tendens för strängens ände att flagna bort, vilket återklingar den observerade "fraying" i verkligt DNA. Att lägga till en korrekt byggsten direkt efter en mismatch kan både påskynda tillväxten och fånga felet på plats, vilket stämmer överens med uppmätta "nästa-nukleotid-effekter" där en efterföljande korrekt nukleotid kan stabilisera ett tidigare misstag. Modellen visar också en avvägning mellan hastighet och noggrannhet: att driva basparningen för svagt eller för starkt försämrar båda fidelity, med ett optimalt mellanting där kopiering är tillräckligt noggrann men inte alltför långsam.
Hur enkel kemi kan skapa varaktig ordning
För en lekmannaläsare är huvudbudskapet att kopiering av genetisk information med hög noggrannhet kanske inte kräver komplexa molekylmaskiner från början. Författarna visar att om korrekta matchningar hjälper tillväxten att fortskrida i en riktning, och om de starka ryggradsbindningarna mellan enheterna bildas tillräckligt snabbt, kan systemet använda samma energi som driver tillväxten för att rensa bort många misstag. I detta perspektiv finslipar och påskyndar enzymer i moderna celler främst en grundläggande fysisk princip snarare än att hitta på den från grunden. Detta erbjuder en plausibel väg för hur tidiga, enzymfria genetiska polymerer skulle kunna ha uppnått tillräckligt hög kopieringsnoggrannhet för att stödja evolution, och illustrerar mer allmänt hur bestående molekylär ordning kan uppstå från enkel, biasad kinetik i en energidriven men enzymlös värld.
Citering: Ghosh, K., Sahu, P., Barik, S. et al. Non-enzymatic error correction in self-replicators without extraneous energy supply. Sci Rep 16, 10165 (2026). https://doi.org/10.1038/s41598-026-40325-9
Nyckelord: livets ursprung, DNA-replikations noggrannhet, icke-enzymatisk replikation, felkorrigering, prebiotisk kemi