Clear Sky Science · pt
Base estrutural da ativação alostérica da isocitrato liase 2 de Mycobacterium tuberculosis
Como uma enzima da tuberculose é ligada como um interruptor molecular
Bactérias da tuberculose sobrevivem dentro de nossos pulmões reaproveitando astutamente seu metabolismo, especialmente quando o açúcar é escasso e apenas gorduras estão disponíveis. Este estudo revela, em detalhe atômico, como uma enzima bacteriana chave chamada isocitrato liase 2 (ICL2) atua como um interruptor molecular que se ativa ao detectar uma pequena molécula transportadora de combustível. Entender esse interruptor não só aprofunda nossa compreensão de como o patógeno se adapta no corpo, mas também aponta maneiras novas e mais sutis de desativar a bactéria com fármacos.
Um atalho de sobrevivência no metabolismo bacteriano
A ICL2 ocupa uma posição central em um atalho metabólico conhecido como via da glicoxilato, que permite que Mycobacterium tuberculosis cresça com fontes de carbono não açucaradas, como ácidos graxos. Essa via é especialmente importante durante a infecção, quando a bactéria vive dentro de células imunes e precisa depender de lipídios derivados do hospedeiro. Ao contrário de muitas enzimas que estão sempre “ligadas”, a ICL2 é regulada por uma pequena molécula chamada acetil-CoA, um transportador central de unidades de carbono produzidas durante a quebra de gorduras. Quando os níveis de acetil-CoA aumentam, a ICL2 torna-se dramaticamente mais ativa, ajudando o patógeno a canalizar carbono pela via da glicoxilato e a sustentar infecções de longa duração.
Dois acréscimos estruturais especiais tornam o interruptor possível
Comparada com enzimas relacionadas, a ICL2 apresenta duas características estruturais incomuns: uma seção helicoidal extra embutida em seu domínio catalítico principal e uma região de cauda separada, chamada domínio C-terminal, pendurada no núcleo por um conector flexível. Os pesquisadores mostraram que a cauda não é necessária para a química básica de quebrar o isocitrato, mas é absolutamente essencial para o grande aumento de atividade desencadeado pelo acetil-CoA. A seção helicoidal, por sua vez, é crítica para manter a enzima montada como um complexo multipartido; quando ela foi removida, a proteína se desfez em unidades simples e perdeu toda atividade detectável, ressaltando que a arquitetura da enzima é inseparável de sua função.
Ligação a longa distância, mudança de forma no núcleo
Para entender como o acetil-CoA ativa a enzima à distância, a equipe usou espectroscopia de ressonância magnética nuclear, cristalografia de raios X, simulações computacionais e mutações direcionadas. Eles focaram no domínio da cauda e descobriram que, por si só, ele geralmente existe como uma única unidade em solução, mas dimeriza — emparelha-se — quando acetil-CoA ou moléculas relacionadas estão presentes. Medições detalhadas mostraram que o acetil-CoA se liga com maior afinidade do que a CoA simples, e que são necessárias concentrações mais altas de CoA para alcançar o mesmo efeito ativador. Isso sugere que a enzima responde de forma sensível aos níveis relativos de diferentes metabólitos baseados em CoA, lendo efetivamente o estado de carbono da célula.
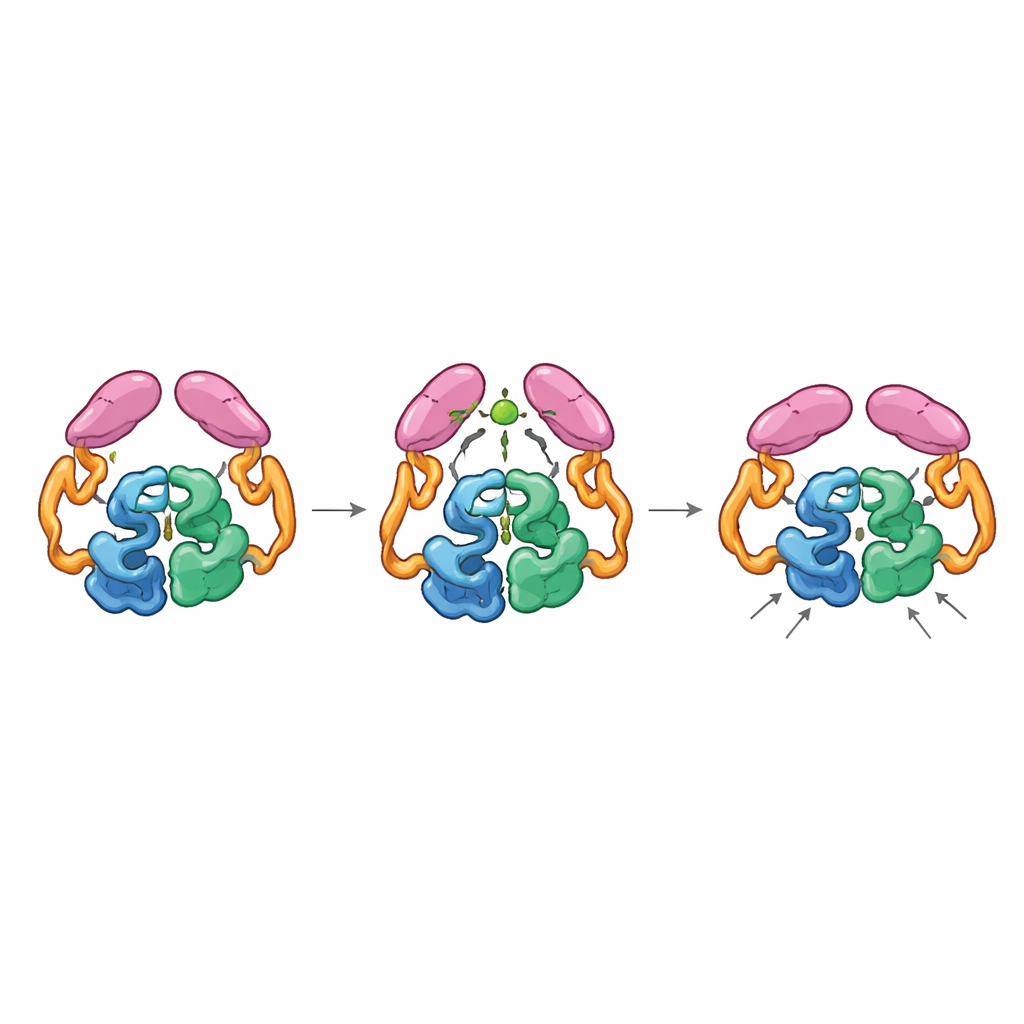
Acoplamento mecânico entre partes distantes
As estruturas da enzima completa em seus estados “desligado” e “ligado” revelaram que os domínios de cauda podem formar pares de duas maneiras distintas, como engrenagens que se encaixam em orientações diferentes. No estado de repouso, as caudas interagem entre si e com a seção helicoidal no núcleo, mantendo a enzima numa disposição menos eficiente em que uma alça chave perto do sítio ativo assume muitas conformações não ideais. Quando o acetil-CoA se liga, ele remodela uma pequena alça na cauda e promove um novo emparelhamento entre as caudas que só é acessível se elas mudarem de posição em relação ao núcleo. Esse movimento rompe contatos anteriores com a região helicoidal e permite que uma alça particular da cauda se encaixe mais perto do centro catalítico, estabilizando a alça do sítio ativo numa conformação mais produtiva. Enzimas mutantes “travadas” na conformação correspondente ao estado ligado ao acetil-CoA foram altamente ativas mesmo sem acetil-CoA, confirmando que é essa forma ativada — e não apenas a presença da pequena molécula — que acelera a reação.
Implicações para tratamentos futuros da tuberculose
Os autores propõem que a ICL2 funcione como um relé sensível a metabólitos: o segmento helicoidal e a cauda mantêm a enzima numa pose inativa até que os níveis de acetil-CoA aumentem, ponto em que a dimerização e o reposicionamento das caudas empurram o sítio ativo para uma configuração de alto desempenho. Como o desenho clássico de inibidores teve dificuldades com enzimas ICL — seus sítios ativos são pequenos e altamente polares — este trabalho destaca novas regiões mais acessíveis para mirar, como os contatos entre a cauda e o segmento helicoidal ou as superfícies que formam o dímero estabilizado por acetil-CoA. Ao perturbar essa fiação alostérica em vez de bloquear a química de forma direta, futuros fármacos podem privar mais efetivamente as bactérias da tuberculose da flexibilidade metabólica de que dependem para persistir no corpo humano.
Citação: Huang, E.YW., Kwai, B.X.C., Jiao, W. et al. Structural basis of allosteric activation of Mycobacterium tuberculosis isocitrate lyase 2. Commun Biol 9, 560 (2026). https://doi.org/10.1038/s42003-026-09821-6
Palavras-chave: metabolismo da tuberculose, regulação alostérica, isocitrato liase 2, sinalização por acetil-CoA, alvos de fármacos antimicrobianos