Clear Sky Science · en
Structural basis of allosteric activation of Mycobacterium tuberculosis isocitrate lyase 2
How a Tuberculosis Enzyme Flips On Like a Molecular Switch
Tuberculosis bacteria survive inside our lungs by cleverly retooling their metabolism, especially when sugar is scarce and only fats are on the menu. This study reveals, in atomic detail, how a key bacterial enzyme called isocitrate lyase 2 (ICL2) acts like a molecular switch that turns on when it senses a small fuel-carrying molecule. Understanding this switch not only deepens our grasp of how the pathogen adapts inside the body, but also points to new, more subtle ways to shut the bacterium down with drugs.
A Survival Shortcut in Bacterial Metabolism
ICL2 sits in the middle of a metabolic shortcut known as the glyoxylate shunt, which lets Mycobacterium tuberculosis grow on non-sugar carbon sources such as fatty acids. This pathway is especially important during infection, when the bacterium lives inside immune cells and must rely on host-derived fats. Unlike many enzymes that are always “on,” ICL2 is tuned by a small molecule called acetyl-CoA, a central carrier of carbon units produced during fat breakdown. When acetyl-CoA levels rise, ICL2 becomes dramatically more active, helping the pathogen funnel carbon through the glyoxylate shunt and sustain long-term infection.
Two Special Structural Add-Ons Make the Switch Possible
Compared with related enzymes, ICL2 carries two unusual structural features: an extra helical section embedded in its main catalytic domain, and a separate tail region, called the C-terminal domain, dangling from the core by a flexible linker. The researchers showed that the tail is not needed for the basic chemistry of breaking down isocitrate, but it is absolutely required for the big boost in activity triggered by acetyl-CoA. The helical section, in turn, is critical for keeping the enzyme assembled into a multi-part complex; when it was removed, the protein fell apart into single units and lost all detectable activity, underscoring that the enzyme’s architecture is inseparable from its function.
Binding Far Away, Changing Shape at the Core
To work out how acetyl-CoA flips the enzyme on from a distance, the team used nuclear magnetic resonance spectroscopy, X-ray crystallography, computer simulations, and targeted mutations. They focused on the tail domain and found that, on its own, it usually exists as a single unit in solution, but dimerizes—pairs up—when acetyl-CoA or related molecules are present. Detailed measurements showed that acetyl-CoA binds tighter than plain CoA, and that higher concentrations of CoA are needed to achieve the same activating effect. This suggests the enzyme responds sensitively to the relative levels of different CoA-based metabolites, effectively reading out the cell’s carbon status.
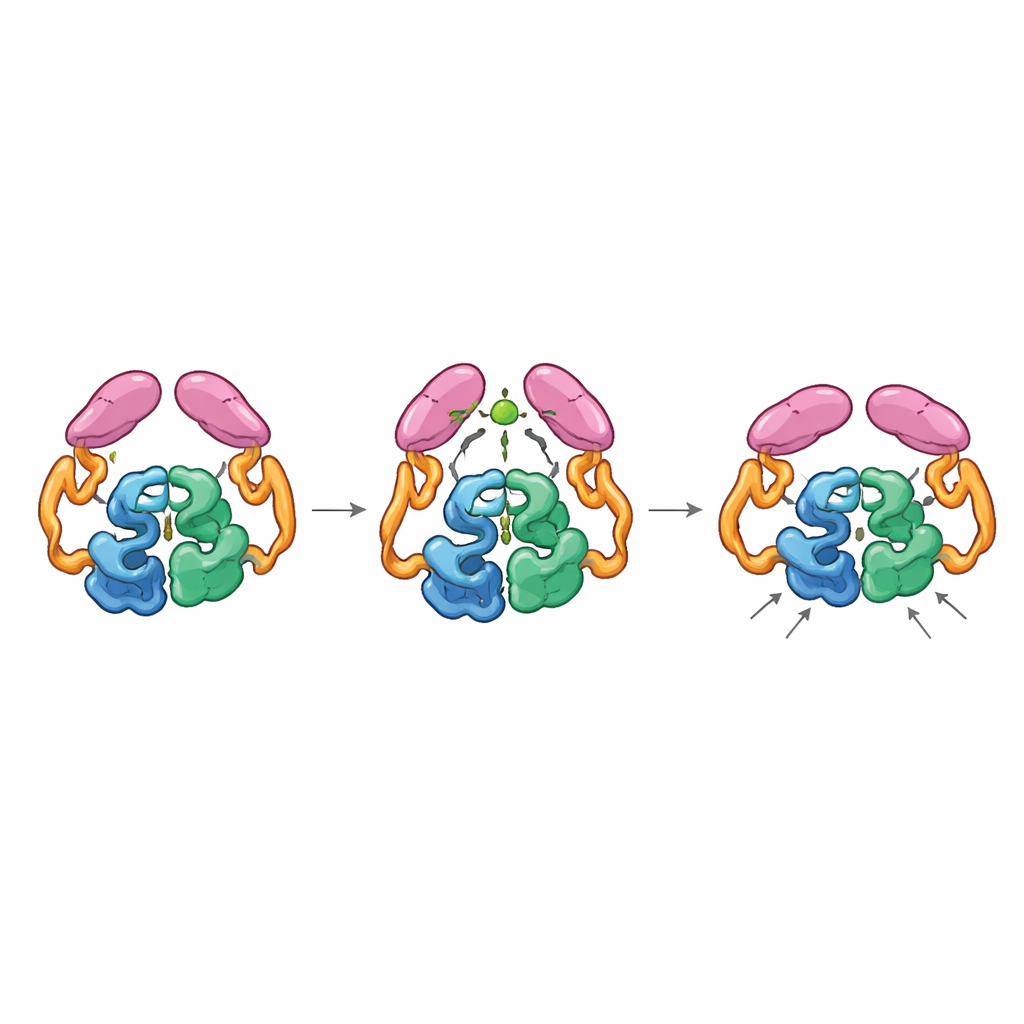
Mechanical Coupling Between Distant Parts
Structures of the full enzyme in its “off” and “on” states revealed that the tail domains can pair up in two distinct ways, like gears that can mesh in different orientations. In the resting state, the tails interact both with each other and with the helical section in the core, holding the enzyme in a less efficient arrangement where a key loop near the active site samples many non-ideal shapes. When acetyl-CoA binds, it reshapes a small loop in the tail and promotes a new pairing between tails that is only accessible if they shift position relative to the core. This movement breaks earlier contacts with the helical region and allows a particular loop on the tail to dock closer to the catalytic center, stabilizing the active-site loop in a more productive conformation. Mutant enzymes “locked” into the acetyl-CoA–bound arrangement were highly active even without acetyl-CoA, confirming that this activated shape, not the presence of the small molecule itself, is what speeds the reaction.
Implications for Future Tuberculosis Treatments
The authors propose that ICL2 acts as a metabolite-sensitive relay: the helical segment and tail hold the enzyme in an inactive pose until acetyl-CoA levels rise, at which point tail dimerization and repositioning push the active site into a high-performance configuration. Because classical inhibitor design has struggled with ICL enzymes—their active sites are small and highly polar—this work highlights new, more accessible regions to target, such as the contacts between the tail and the helical segment or the surfaces that form the acetyl-CoA–stabilized dimer. By disrupting this allosteric wiring rather than blocking chemistry outright, future drugs might more effectively starve tuberculosis bacteria of the metabolic flexibility they need to persist in the human body.
Citation: Huang, E.YW., Kwai, B.X.C., Jiao, W. et al. Structural basis of allosteric activation of Mycobacterium tuberculosis isocitrate lyase 2. Commun Biol 9, 560 (2026). https://doi.org/10.1038/s42003-026-09821-6
Keywords: tuberculosis metabolism, allosteric regulation, isocitrate lyase 2, acetyl-CoA signaling, antimicrobial drug targets