Clear Sky Science · it
Base strutturale dell’attivazione allosterica dell’isocitrato liasi 2 di Mycobacterium tuberculosis
Come un enzima della tubercolosi si accende come un interruttore molecolare
I batteri della tubercolosi sopravvivono nei nostri polmoni rimodellando abilmente il loro metabolismo, soprattutto quando lo zucchero scarseggia e l’unica fonte disponibile sono i grassi. Questo studio rivela, a livello atomico, come un enzima chiave del patogeno chiamato isocitrato liasi 2 (ICL2) si comporti come un interruttore molecolare che si attiva quando rileva una piccola molecola trasportatrice di carbonio. Capire questo interruttore non solo approfondisce la nostra comprensione di come il patogeno si adatti all’interno dell’organismo, ma indica anche nuovi modi più sottili per bloccare il batterio con farmaci.
Una scorciatoia di sopravvivenza nel metabolismo batterico
ICL2 si trova al centro di una scorciatoia metabolica nota come via del gioxilato, che permette a Mycobacterium tuberculosis di crescere su fonti di carbonio non zuccherine come gli acidi grassi. Questa via è particolarmente importante durante l’infezione, quando il batterio vive all’interno delle cellule immunitarie e deve fare affidamento sui lipidi derivati dall’ospite. Diversamente da molti enzimi costantemente “attivi”, ICL2 è regolata da una piccola molecola chiamata acetil-CoA, un vettore centrale di unità di carbonio prodotte durante la degradazione dei grassi. Quando i livelli di acetil-CoA aumentano, ICL2 diventa dramaticamente più attiva, aiutando il patogeno a convogliare il carbonio attraverso la via del gioxilato e a sostenere infezioni di lunga durata.
Due inserti strutturali speciali rendono possibile l’interruttore
Rispetto agli enzimi correlati, ICL2 presenta due caratteristiche strutturali insolite: una sezione elicoidale aggiuntiva inserita nel suo dominio catalitico principale e una regione terminale separata, chiamata dominio C-terminale, sospesa dal nucleo tramite un legante flessibile. I ricercatori hanno dimostrato che la coda non è necessaria per la chimica di base della degradazione dell’isocitrato, ma è assolutamente richiesta per il forte aumento di attività innescato dall’acetil-CoA. La sezione elicoidale, a sua volta, è cruciale per mantenere l’enzima assemblato in un complesso multiparto; quando è stata rimossa, la proteina si disgregava in unità singole e perdeva ogni attività rilevabile, sottolineando come l’architettura dell’enzima sia inseparabile dalla sua funzione.
Legame a distanza, cambiamento di forma al centro
Per capire come l’acetil-CoA attivi l’enzima a distanza, il team ha usato spettroscopia di risonanza magnetica nucleare, cristallografia a raggi X, simulazioni al computer e mutazioni mirate. Si sono concentrati sul dominio a coda e hanno scoperto che, da solo, esso in soluzione esiste di solito come monomero, ma dimerizza — si accoppia in coppie — quando sono presenti acetil-CoA o molecole affini. Misurazioni dettagliate hanno mostrato che l’acetil-CoA si lega con maggiore affinità rispetto al solo CoA e che sono necessarie concentrazioni più alte di CoA per ottenere lo stesso effetto attivante. Questo suggerisce che l’enzima risponda in modo sensibile ai livelli relativi dei diversi metaboliti legati al CoA, leggendo di fatto lo stato carbonioso della cellula.
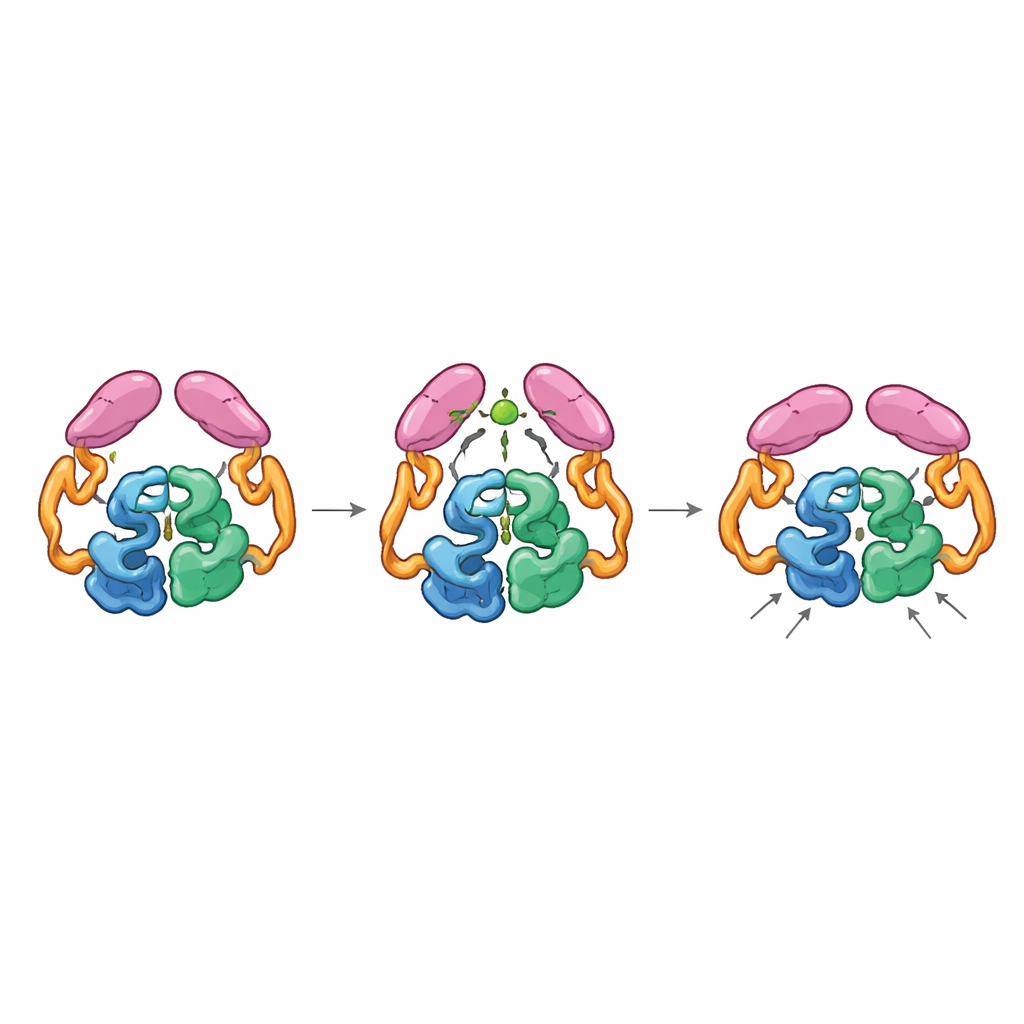
Accoppiamento meccanico tra parti distanti
Le strutture dell’enzima completo nei suoi stati “spento” e “acceso” hanno rivelato che i domini a coda possono accoppiarsi in due modi distinti, come ingranaggi che possono innestarsi in orientamenti diversi. Nello stato di riposo, le code interagiscono sia tra loro sia con la sezione elicoidale del nucleo, mantenendo l’enzima in un assetto meno efficiente in cui un anello chiave vicino al sito attivo esplora molte conformazioni non ideali. Quando si lega l’acetil-CoA, questo rimodella un piccolo anello nella coda e favorisce un nuovo accoppiamento tra le code accessibile solo se esse si spostano rispetto al nucleo. Questo movimento rompe i contatti precedenti con la regione elicoidale e permette a un particolare anello della coda di posizionarsi più vicino al centro catalitico, stabilizzando l’anello del sito attivo in una conformazione più produttiva. Enzimi mutanti “bloccati” nella disposizione tipica del complesso con acetil-CoA erano altamente attivi anche in assenza di acetil-CoA, confermando che è la forma attivata, non la mera presenza della piccola molecola, a velocizzare la reazione.
Implicazioni per futuri trattamenti della tubercolosi
Gli autori propongono che ICL2 funzioni come un relè sensibile ai metaboliti: il segmento elicoidale e la coda mantengono l’enzima in una posa inattiva finché i livelli di acetil-CoA non aumentano, momento in cui la dimerizzazione e il riposizionamento delle code spingono il sito attivo in una configurazione ad alte prestazioni. Poiché la progettazione classica di inibitori ha avuto difficoltà con gli enzimi ICL — i loro siti attivi sono piccoli e altamente polari — questo lavoro mette in luce nuove regioni più accessibili da colpire, come i contatti tra coda e segmento elicoidale o le superfici che formano il dimero stabilizzato dall’acetil-CoA. Disgregando questo cablaggio allosterico piuttosto che bloccando direttamente la chimica, i farmaci futuri potrebbero essere in grado di privare in modo più efficace i batteri della tubercolosi della flessibilità metabolica necessaria per persistere nell’organismo umano.
Citazione: Huang, E.YW., Kwai, B.X.C., Jiao, W. et al. Structural basis of allosteric activation of Mycobacterium tuberculosis isocitrate lyase 2. Commun Biol 9, 560 (2026). https://doi.org/10.1038/s42003-026-09821-6
Parole chiave: metabolismo della tubercolosi, regolazione allosterica, isocitrato liasi 2, segnalizzazione dell'acetil-CoA, bersagli farmacologici antimicrobici