Clear Sky Science · pl
Strukturalne podstawy allosterycznej aktywacji izocytrynianowej liazy 2 Mycobacterium tuberculosis
Jak enzym prątków gruźlicy przełącza się jak molekularny wyłącznik
Bakterie wywołujące gruźlicę przetrwają w płucach, sprytnie przeorganizowując swój metabolizm, zwłaszcza gdy brakuje cukrów i dostępne są jedynie tłuszcze. W tym badaniu ukazano, w skali atomowej, w jaki sposób kluczowy enzym bakteryjny zwany izocytrynianową liazą 2 (ICL2) działa jak molekularny przełącznik, który włącza się po wykryciu małej cząsteczki przenoszącej paliwo. Zrozumienie tego mechanizmu pogłębia naszą wiedzę o tym, jak patogen adaptuje się w organizmie, i wskazuje nowe, subtelniejsze strategie jego unieruchomienia za pomocą leków.
Skrót przetrwania w metabolizmie bakterii
ICL2 znajduje się w centrum metabolicznego skrótu znanego jako szlak gloksylatowy, który pozwala Mycobacterium tuberculosis rosnąć na źródłach węgla innych niż cukry, takich jak kwasy tłuszczowe. Ta droga jest szczególnie ważna podczas infekcji, gdy bakteria żyje wewnątrz komórek układu odpornościowego i musi polegać na tłuszczach pochodzących od gospodarza. W przeciwieństwie do wielu enzymów, które są stale „włączone”, aktywność ICL2 regulowana jest przez małą cząsteczkę — acetylo-CoA, centralnego nośnika jednostek węglowych produkowanych podczas rozkładu tłuszczów. Gdy poziom acetylo-CoA rośnie, aktywność ICL2 zwiększa się znacznie, pomagając patogenowi skierować węgiel przez szlak gloksylatowy i podtrzymać długotrwałą infekcję.
Dwa specjalne dodatki strukturalne umożliwiają przełączenie
W porównaniu z pokrewnymi enzymami, ICL2 ma dwie nietypowe cechy strukturalne: dodatkową sekcję helikalną osadzoną w głównej domenie katalitycznej oraz oddzielny odcinek ogonowy, zwany domeną C-terminalną, zwisający z rdzenia na elastycznym łączniku. Badacze wykazali, że ogon nie jest potrzebny do podstawowej chemii rozkładu izocytrynianu, ale jest absolutnie niezbędny do silnego wzrostu aktywności wywołanego przez acetylo-CoA. Sekcja helikalna z kolei jest krytyczna dla utrzymania enzymu w złożonym wieloczęściowym układzie; po jej usunięciu białko rozpadało się na pojedyncze jednostki i traciło wykrywalną aktywność, co podkreśla, że architektura enzymu jest nierozerwalna z jego funkcją.
Wiązanie daleko, zmiana kształtu w centrum
Aby wyjaśnić, jak acetylo-CoA włącza enzym z daleka, zespół zastosował spektroskopię rezonansu magnetycznego jądrowego, krystalografię promieniowania rentgenowskiego, symulacje komputerowe oraz ukierunkowane mutacje. Skupili się na domenie ogonowej i odkryli, że sama w roztworze zwykle występuje jako monomer, ale dimeryzuje — tworzy pary — gdy obecne są acetylo-CoA lub pokrewne cząsteczki. Szczegółowe pomiary wykazały, że acetylo-CoA wiąże się silniej niż sam CoA, a do osiągnięcia tego samego efektu aktywującego potrzeba wyższych stężeń CoA. Sugeruje to, że enzym reaguje wrażliwie na względne poziomy różnych metabolitów opartych na CoA, efektywnie odczytując stan węglowy komórki.
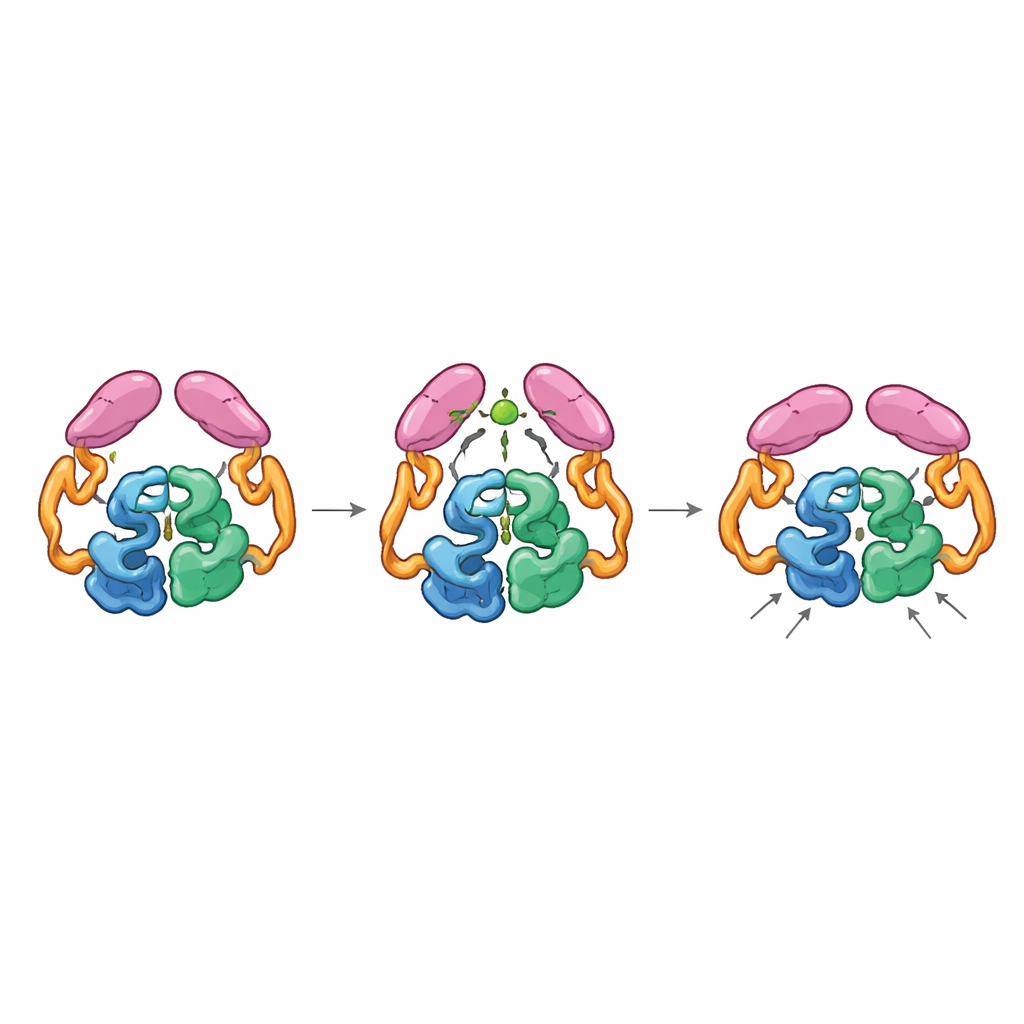
Sprzężenie mechaniczne między odległymi częściami
Struktury pełnego enzymu w stanach „wyłączonym” i „włączonym” ujawniły, że domeny ogonowe mogą parować się na dwa różne sposoby, jak zębatki mogą zazębiać się w różnych orientacjach. W stanie spoczynkowym ogony oddziałują zarówno ze sobą, jak i z sekcją helikalną w rdzeniu, utrzymując enzym w mniej wydajnym ustawieniu, gdzie kluczowa pętla w pobliżu centrum aktywnego przyjmuje wiele nieoptymalnych kształtów. Gdy acetylo-CoA wiąże się, przekształca małą pętlę w ogonie i sprzyja nowemu parowaniu ogonów, które jest dostępne tylko jeśli przesuną one pozycję względem rdzenia. Ten ruch przerywa wcześniejsze kontakty z regionem helikalnym i pozwala konkretnej pętli ogona zakotwiczyć się bliżej centrum katalitycznego, stabilizując pętlę miejsca aktywnego w bardziej produktywnej konformacji. Mutanty enzymu „zablokowane” w układzie odpowiadającym wiązaniu acetylo-CoA były wysoce aktywne nawet bez obecności acetylo-CoA, potwierdzając, że to aktywowany kształt, a nie sama obecność małej cząsteczki, przyspiesza reakcję.
Implikacje dla przyszłych terapii gruźlicy
Autorzy proponują, że ICL2 działa jak przekaźnik czuły na metabolity: segment helikalny i ogon utrzymują enzym w nieaktywniej pozie, dopóki poziom acetylo-CoA nie wzrośnie, po czym dimeryzacja i repozycjonowanie ogonów popychają miejsce aktywne do wydajnej konfiguracji. Ponieważ klasyczne projektowanie inhibitorów miało trudności z enzymami ICL — ich miejsca aktywne są małe i wysoce polarne — ta praca uwydatnia nowe, bardziej dostępne obszary do ataku, takie jak kontakty między ogonem a segmentem helikalnym lub powierzchnie tworzące dimer stabilizowany przez acetylo-CoA. Zakłócając to allosteryczne okablowanie zamiast blokowania samej chemii, przyszłe leki mogłyby skuteczniej pozbawić prątki gruźlicy elastyczności metabolicznej, której potrzebują do przetrwania w organizmie człowieka.
Cytowanie: Huang, E.YW., Kwai, B.X.C., Jiao, W. et al. Structural basis of allosteric activation of Mycobacterium tuberculosis isocitrate lyase 2. Commun Biol 9, 560 (2026). https://doi.org/10.1038/s42003-026-09821-6
Słowa kluczowe: metabolizm gruźlicy, regulacja allosteryczna, izocytrynianowa liaza 2, sygnalizacja acetylo-CoA, cele leków przeciwmikrobiowych