Clear Sky Science · fr
Base structurale de l’activation allostérique de l’isocitrate lyase 2 de Mycobacterium tuberculosis
Comment une enzyme de la tuberculose s’enclenche comme un interrupteur moléculaire
Les bactéries responsables de la tuberculose survivent dans nos poumons en réorientant astucieusement leur métabolisme, notamment lorsque le glucose se fait rare et que seuls les lipides sont disponibles. Cette étude révèle, au niveau atomique, comment une enzyme bactérienne clé appelée isocitrate lyase 2 (ICL2) fonctionne comme un interrupteur moléculaire qui s’active lorsqu’elle détecte une petite molécule porteuse d’énergie. Comprendre cet interrupteur approfondit non seulement notre connaissance des adaptations du pathogène dans l’organisme, mais indique aussi de nouvelles voies plus subtiles pour l’affaiblir par des médicaments.
Un raccourci de survie dans le métabolisme bactérien
ICL2 se situe au cœur d’un raccourci métabolique connu sous le nom de shunt du glyoxylate, qui permet à Mycobacterium tuberculosis de croître sur des sources de carbone non glucidiques telles que les acides gras. Cette voie est particulièrement importante durant l’infection, lorsque la bactérie vit à l’intérieur des cellules immunitaires et doit s’appuyer sur les lipides provenant de l’hôte. Contrairement à de nombreuses enzymes toujours « actives », ICL2 est modulée par une petite molécule, l’acétyl-CoA, un vecteur central d’unités carbonées produit lors de la dégradation des graisses. Quand les niveaux d’acétyl-CoA augmentent, ICL2 voit son activité augmenter fortement, aidant le pathogène à canaliser le carbone via le shunt du glyoxylate et à maintenir une infection chronique.
Deux ajouts structurels particuliers rendent l’interrupteur possible
Comparée à des enzymes apparentées, ICL2 présente deux caractéristiques structurales inhabituelles : une section hélicoïdale supplémentaire intégrée dans son domaine catalytique principal, et une région en queue distincte, appelée domaine C-terminal, pendant depuis le noyau via une liaison flexible. Les chercheurs ont montré que la queue n’est pas nécessaire pour la chimie de base de la dégradation de l’isocitrate, mais qu’elle est absolument requise pour le fort gain d’activité déclenché par l’acétyl-CoA. La section hélicoïdale, elle, est cruciale pour maintenir l’enzyme assemblée en un complexe multiprotéique ; lorsqu’elle a été supprimée, la protéine s’est dissociée en unités monomériques et a perdu toute activité détectable, soulignant que l’architecture de l’enzyme est indissociable de sa fonction.
Fixation à distance, changement de conformation au cœur
Pour élucider comment l’acétyl-CoA active l’enzyme à distance, l’équipe a utilisé la spectroscopie de résonance magnétique nucléaire, la cristallographie aux rayons X, des simulations informatiques et des mutations ciblées. Ils se sont concentrés sur le domaine de la queue et ont constaté que, isolée, elle existe généralement en solution sous forme monomérique, mais qu’elle se dimérise — se couple en paires — en présence d’acétyl-CoA ou de molécules apparentées. Des mesures détaillées ont montré que l’acétyl-CoA se lie plus fortement que la CoA simple, et que des concentrations plus élevées de CoA sont nécessaires pour obtenir le même effet activateur. Cela suggère que l’enzyme répond finement aux niveaux relatifs des différents métabolites liés à la CoA, lisant en quelque sorte l’état carboné de la cellule.
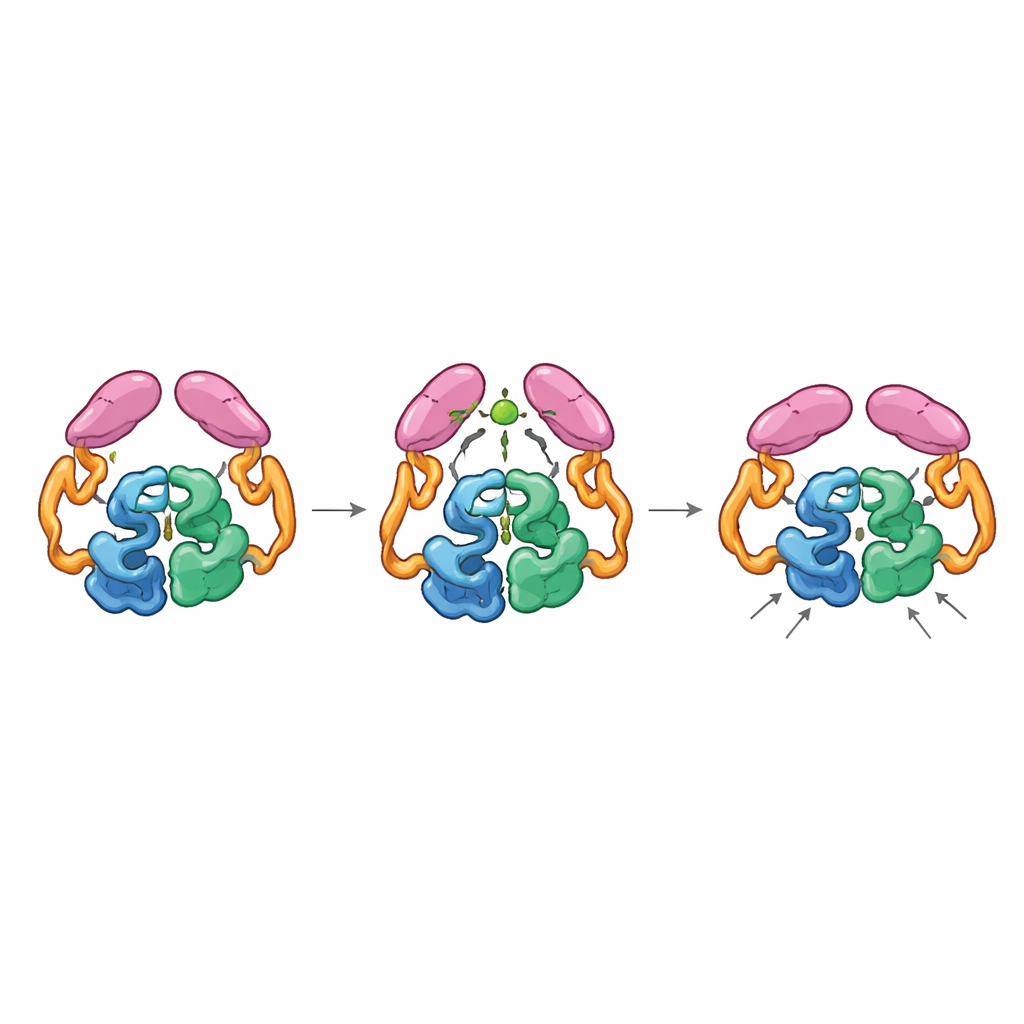
Couplage mécanique entre parties éloignées
Les structures de l’enzyme complète dans ses états « éteint » et « allumé » ont révélé que les domaines en queue peuvent s’apparier de deux manières distinctes, comme des engrenages pouvant s’emboîter dans différentes orientations. À l’état de repos, les queues interagissent entre elles et avec la section hélicoïdale du noyau, maintenant l’enzyme dans une configuration moins efficace où une boucle clé près du site actif explore de nombreuses conformations non optimales. Lorsque l’acétyl-CoA se lie, il reconfigure une petite boucle de la queue et favorise un nouvel appariement des queues qui n’est accessible que si elles déplacent leur position par rapport au noyau. Ce mouvement rompt les contacts antérieurs avec la région hélicoïdale et permet à une boucle particulière de la queue de se rapprocher du centre catalytique, stabilisant la boucle du site actif dans une conformation plus productive. Des enzymes mutantes « verrouillées » dans la configuration liée à l’acétyl-CoA étaient hautement actives même sans acétyl-CoA, confirmant que c’est cette conformation activée — et non la simple présence de la petite molécule — qui accélère la réaction.
Conséquences pour les traitements futurs de la tuberculose
Les auteurs proposent qu’ICL2 agisse comme un relais sensible aux métabolites : le segment hélicoïdal et la queue maintiennent l’enzyme dans une pose inactive jusqu’à ce que les niveaux d’acétyl-CoA augmentent, moment auquel la dimérisation et le repositionnement des queues poussent le site actif vers une configuration haute performance. Parce que la conception d’inhibiteurs classiques a buté contre les enzymes ICL — leurs sites actifs sont petits et très polaires — ce travail met en lumière de nouvelles régions plus accessibles à cibler, telles que les contacts entre la queue et le segment hélicoïdal ou les surfaces qui forment le dimère stabilisé par l’acétyl-CoA. En perturbant ce câblage allostérique plutôt qu’en bloquant directement la chimie, de futurs médicaments pourraient affamer plus efficacement les bactéries tuberculeuses de la flexibilité métabolique dont elles ont besoin pour persister dans l’organisme humain.
Citation: Huang, E.YW., Kwai, B.X.C., Jiao, W. et al. Structural basis of allosteric activation of Mycobacterium tuberculosis isocitrate lyase 2. Commun Biol 9, 560 (2026). https://doi.org/10.1038/s42003-026-09821-6
Mots-clés: métabolisme de la tuberculose, régulation allostérique, isocitrate lyase 2, signalisation par l’acétyl-CoA, cibles pharmacologiques antimicrobiennes