Clear Sky Science · de
Strukturelle Grundlage der allosterischen Aktivierung der Isocitrat-Lyase 2 von Mycobacterium tuberculosis
Wie ein Tuberkulose‑Enzym wie ein molekularer Schalter umspringt
Tuberkulosebakterien überleben in unseren Lungen, indem sie ihren Stoffwechsel geschickt umlenken, vor allem wenn Zucker knapp ist und nur Fette verfügbar sind. Diese Studie zeigt auf atomarer Ebene, wie ein zentrales bakterielles Enzym, die Isocitrat‑Lyase 2 (ICL2), wie ein molekularer Schalter funktioniert, der einschaltet, wenn es ein kleines, energieübertragendes Molekül erkennt. Das Verständnis dieses Schalters vertieft nicht nur unser Wissen darüber, wie der Erreger sich im Körper anpasst, sondern weist auch auf neue, subtilere Wege hin, das Bakterium mit Arzneistoffen außer Gefecht zu setzen.
Ein Überlebens‑Kurzschluss im bakteriellen Stoffwechsel
ICL2 sitzt in der Mitte eines stoffwechselbiologischen Kurzschlusses, bekannt als Glykoxylat-Shunt, der es Mycobacterium tuberculosis erlaubt, auf nicht‑zuckerhaltigen Kohlenstoffquellen wie Fettsäuren zu wachsen. Dieser Weg ist besonders wichtig während der Infektion, wenn das Bakterium in Immunzellen lebt und auf vom Wirt bereitgestellte Fette angewiesen ist. Anders als viele Enzyme, die ständig »an« sind, wird ICL2 durch ein kleines Molekül namens Acetyl‑CoA moduliert, einen zentralen Kohlenstoffüberträger, der bei der Fettspaltung entsteht. Bei steigenden Acetyl‑CoA‑Spiegeln wird ICL2 deutlich aktiver und hilft dem Erreger, Kohlenstoff durch den Glykoxylat‑Shunt zu leiten und so eine langanhaltende Infektion aufrechtzuerhalten.
Zwei spezielle strukturelle Aufsätze ermöglichen den Schalter
Im Vergleich zu verwandten Enzymen trägt ICL2 zwei ungewöhnliche strukturelle Merkmale: einen zusätzlichen Helixabschnitt, eingebettet in seine katalytische Hauptdomäne, und eine separate Schwanzregion, die C‑terminale Domäne, die durch eine flexible Verbindung vom Kern herabhängt. Die Forschenden zeigten, dass der Schwanz für die eigentliche Chemie des Isocitratabbaus nicht nötig ist, wohl aber unbedingt erforderlich für die starke Aktivitätssteigerung, die durch Acetyl‑CoA ausgelöst wird. Der Helixabschnitt ist wiederum entscheidend, um das Enzym als Mehrteilkomplex zusammenzuhalten; wurde er entfernt, zerfiel das Protein in Einzelheiten und verlor jegliche nachweisbare Aktivität, was unterstreicht, dass die Architektur des Enzyms untrennbar mit seiner Funktion verbunden ist.
Bindung in der Ferne, Formänderung im Kern
Um herauszufinden, wie Acetyl‑CoA das Enzym aus der Distanz einschaltet, nutzte das Team Kernspinresonanzspektroskopie, Röntgenkristallographie, Computersimulationen und gezielte Mutationen. Der Fokus lag auf der Schwanzdomäne: Isoliert liegt sie meist als Einzelstück in Lösung vor, dimerisiert jedoch — paarweise —, wenn Acetyl‑CoA oder verwandte Moleküle vorhanden sind. Detaillierte Messungen zeigten, dass Acetyl‑CoA stärker bindet als freies CoA und dass höhere CoA‑Konzentrationen nötig sind, um denselben aktivierenden Effekt zu erzielen. Das legt nahe, dass das Enzym empfindlich auf die relativen Spiegel unterschiedlicher CoA‑basierter Metaboliten reagiert und damit effektiv den Kohlenstoffstatus der Zelle abliest.
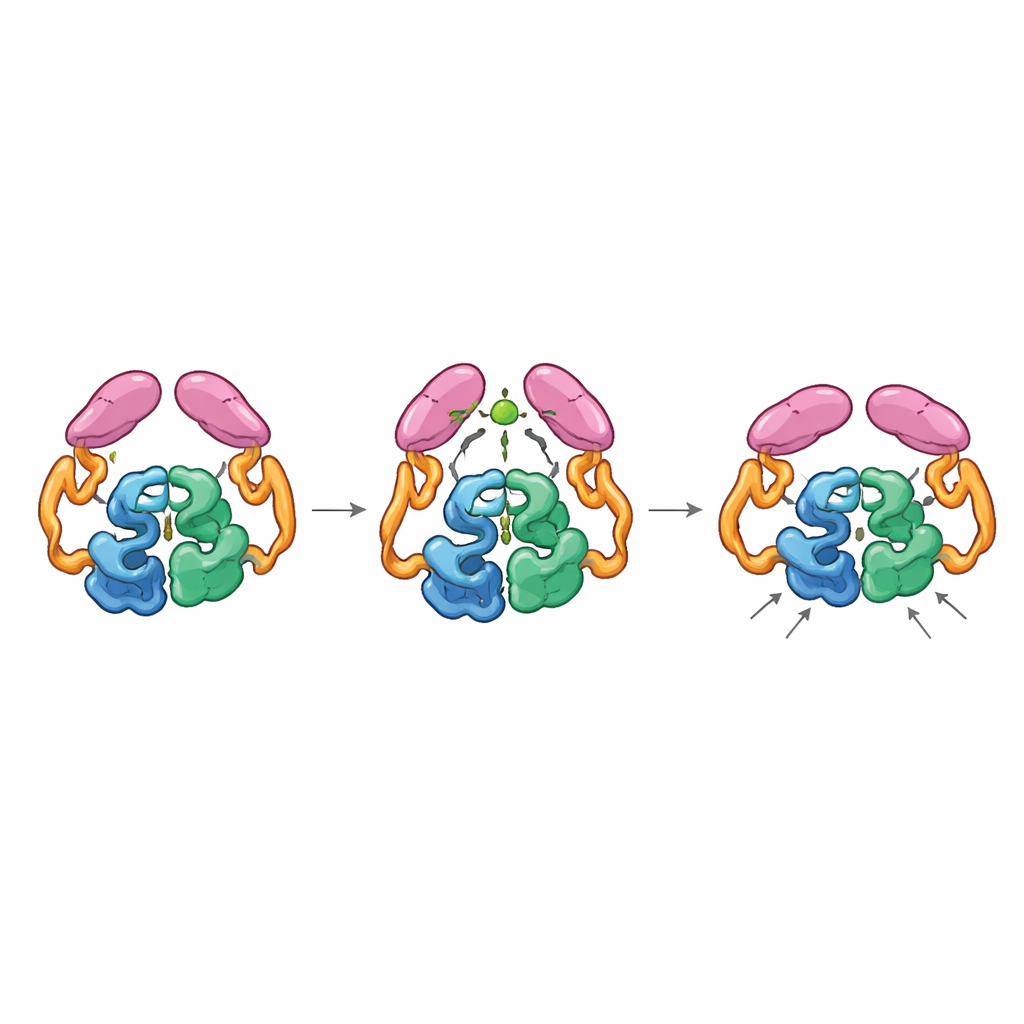
Mechanische Kopplung zwischen weit entfernten Teilen
Strukturen des vollständigen Enzyms in seinen »aus«‑ und »an«‑Zuständen zeigten, dass die Schwanzdomänen in zwei verschiedenen Weisen paaren können, ähnlich wie Zahnräder, die in unterschiedlichen Orientierungen ineinander greifen. Im Ruhezustand interagieren die Schwänze sowohl miteinander als auch mit dem Helixabschnitt im Kern und halten das Enzym in einer weniger effizienten Anordnung, in der eine Schlüssel‑Schleife nahe der aktiven Stelle viele nicht optimale Formen einnimmt. Wenn Acetyl‑CoA bindet, formt es eine kleine Schleife im Schwanz um und fördert eine neue Paarung der Schwänze, die nur zugänglich ist, wenn sie sich gegenüber dem Kern verschieben. Diese Bewegung löst frühere Kontakte zur Helixregion und erlaubt einer bestimmten Schleife am Schwanz, näher an das katalytische Zentrum anzudocken, wodurch die Schleife in der aktiven Stelle in einer produktiveren Konformation stabilisiert wird. Mutanten, die in der Acetyl‑CoA‑gebundenen Anordnung »verriegelt« waren, zeigten hohe Aktivität auch ohne Acetyl‑CoA, was bestätigt, dass diese aktivierte Form — nicht die bloße Anwesenheit des kleinen Moleküls — die Reaktion beschleunigt.
Folgen für künftige Tuberkulose‑Behandlungen
Die Autorinnen und Autoren schlagen vor, dass ICL2 als ein metabolitsensitiver Vermittler fungiert: Der Helix‑Abschnitt und der Schwanz halten das Enzym in einer inaktiven Pose, bis die Acetyl‑CoA‑Spiegel steigen; dann treiben Schwanzdimerisierung und Umlagerung die aktive Stelle in eine leistungsfähige Konfiguration. Da klassische Inhibitor‑Designs bei ICL‑Enzymen schwer erfolgreich waren — ihre aktiven Stellen sind klein und stark polar — hebt diese Arbeit neue, besser zugängliche Bereiche als Angriffspunkte hervor, etwa die Kontaktflächen zwischen Schwanz und Helixabschnitt oder die Oberflächen, die das durch Acetyl‑CoA stabilisierte Dimer bilden. Durch die Störung dieser allosterischen Verschaltung statt des direkten Blockierens der Chemie könnten künftige Wirkstoffe Mykobakterien effektiver die metabolische Flexibilität nehmen, die sie benötigen, um im menschlichen Körper zu bestehen.
Zitation: Huang, E.YW., Kwai, B.X.C., Jiao, W. et al. Structural basis of allosteric activation of Mycobacterium tuberculosis isocitrate lyase 2. Commun Biol 9, 560 (2026). https://doi.org/10.1038/s42003-026-09821-6
Schlüsselwörter: Tuberkulose-Stoffwechsel, allosterische Regulation, Isocitrat-Lyase 2, Acetyl‑CoA‑Signalgebung, Ziele für antimikrobielle Wirkstoffe