Clear Sky Science · es
Base estructural de la activación alostérica de la isocitrato liasa 2 de Mycobacterium tuberculosis
Cómo una enzima de la tuberculosis se enciende como un interruptor molecular
Las bacterias de la tuberculosis sobreviven en nuestros pulmones reconfigurando su metabolismo con ingenio, sobre todo cuando el azúcar escasea y sólo hay lípidos disponibles. Este estudio revela, con detalle atómico, cómo una enzima bacteriana clave llamada isocitrato liasa 2 (ICL2) funciona como un interruptor molecular que se activa al detectar una pequeña molécula portadora de combustible. Comprender este interruptor no solo profundiza nuestro conocimiento de cómo el patógeno se adapta dentro del organismo, sino que también señala formas nuevas y más sutiles de neutralizarlo con fármacos.
Un atajo de supervivencia en el metabolismo bacteriano
ICL2 ocupa un lugar central en un atajo metabólico conocido como la derivación del glioxilato, que permite a Mycobacterium tuberculosis crecer con fuentes de carbono que no son azúcares, como los ácidos grasos. Esta vía es especialmente importante durante la infección, cuando la bacteria vive dentro de las células inmunitarias y debe depender de los lípidos procedentes del hospedador. A diferencia de muchas enzimas que están siempre “encendidas”, ICL2 está regulada por una pequeña molécula llamada acetil-CoA, un transportador central de unidades de carbono producido durante la degradación de grasas. Cuando aumentan los niveles de acetil-CoA, ICL2 se vuelve mucho más activa, ayudando al patógeno a canalizar carbono a través de la derivación del glioxilato y a sostener la infección a largo plazo.
Dos estructuras añadidas especiales hacen posible el interruptor
En comparación con enzimas afines, ICL2 presenta dos características estructurales inusuales: una sección helicoidal extra incrustada en su dominio catalítico principal y una cola separada, denominada dominio C-terminal, que cuelga del núcleo mediante un enlace flexible. Los investigadores demostraron que la cola no es necesaria para la química básica de romper el isocitrato, pero sí es absolutamente imprescindible para el gran aumento de actividad desencadenado por el acetil-CoA. La sección helicoidal, a su vez, es crítica para mantener la enzima ensamblada en un complejo multipartito; cuando se eliminó, la proteína se desensambló en unidades monoméricas y perdió toda actividad detectable, lo que subraya que la arquitectura de la enzima es inseparable de su función.
Unión a distancia, cambio de forma en el núcleo
Para dilucidar cómo el acetil-CoA activa la enzima desde la distancia, el equipo usó espectroscopía de resonancia magnética nuclear, cristalografía de rayos X, simulaciones por ordenador y mutagénesis dirigida. Se centraron en el dominio de la cola y encontraron que, por sí solo, suele existir como monómero en solución, pero se dimeriza —se empareja— cuando están presentes acetil-CoA u otras moléculas relacionadas. Mediciones detalladas mostraron que el acetil-CoA se une con mayor afinidad que la CoA simple, y que se requieren concentraciones más altas de CoA para lograr el mismo efecto activador. Esto sugiere que la enzima responde con sensibilidad a los niveles relativos de distintos metabolitos basados en CoA, interpretando efectivamente el estado de carbono de la célula.
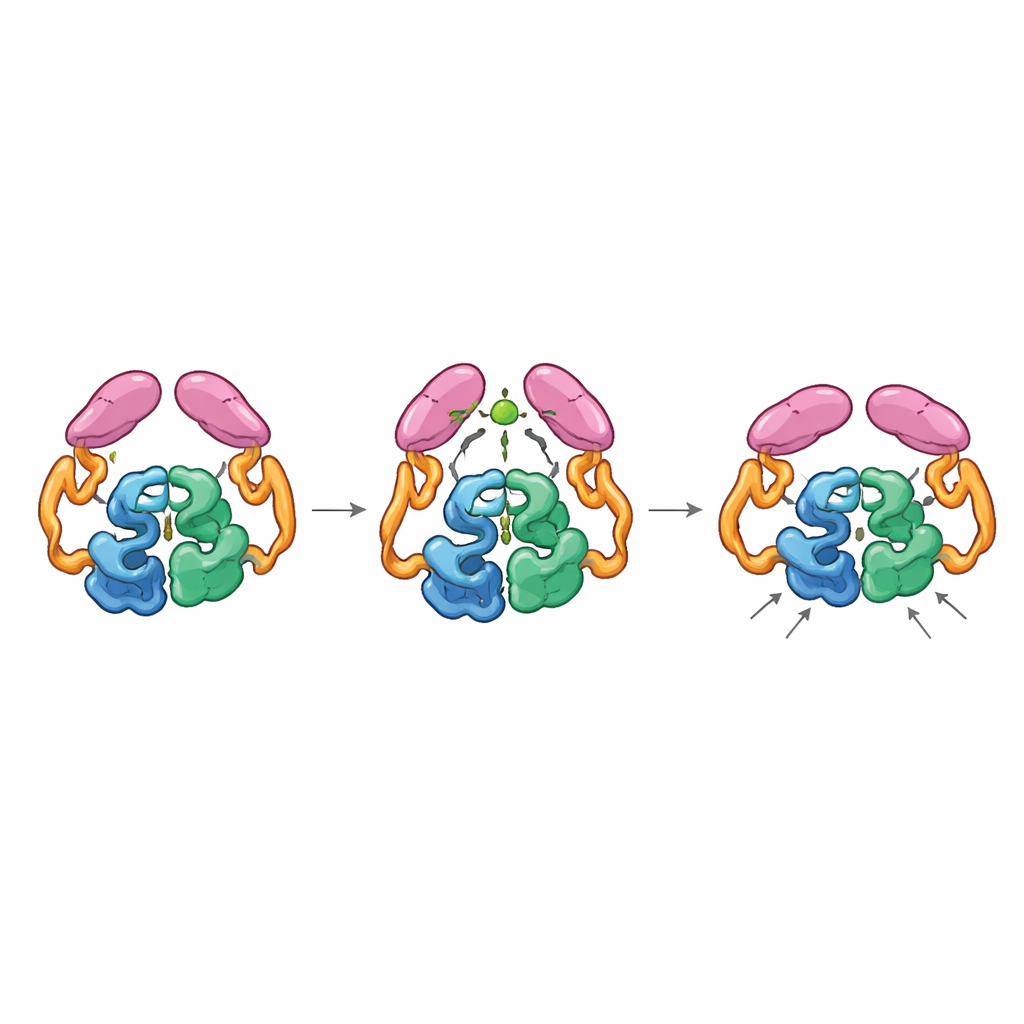
Acoplamiento mecánico entre partes distantes
Las estructuras de la enzima completa en sus estados “apagado” y “encendido” revelaron que los dominios de la cola pueden emparejarse de dos maneras distintas, como engranajes que pueden engranar en diferentes orientaciones. En el estado de reposo, las colas interactúan tanto entre sí como con la sección helicoidal del núcleo, manteniendo la enzima en una disposición menos eficiente donde un bucle clave cercano al sitio activo explora muchas conformaciones no ideales. Cuando se une el acetil-CoA, remodela un pequeño bucle en la cola y promueve un nuevo emparejamiento entre colas que solo es accesible si éstas cambian de posición respecto al núcleo. Este movimiento rompe contactos previos con la región helicoidal y permite que un bucle particular de la cola se acople más cerca del centro catalítico, estabilizando el bucle del sitio activo en una conformación más productiva. Enzimas mutantes “bloqueadas” en la disposición propia del acetil-CoA mostraron una alta actividad incluso sin acetil-CoA, lo que confirma que es esa forma activada, y no la presencia de la pequeña molécula en sí, la que acelera la reacción.
Implicaciones para futuros tratamientos contra la tuberculosis
Los autores proponen que ICL2 actúa como un relevo sensible a metabolitos: el segmento helicoidal y la cola mantienen la enzima en una pose inactiva hasta que suben los niveles de acetil-CoA; en ese punto, la dimerización y el reposicionamiento de las colas empujan el sitio activo hacia una configuración de alto rendimiento. Dado que el diseño clásico de inhibidores ha tropezado con las ICL —sus sitios activos son pequeños y muy polares—, este trabajo destaca nuevas regiones más accesibles para atacar, como los contactos entre la cola y el segmento helicoidal o las superficies que forman el dímero estabilizado por acetil-CoA. Al interrumpir este cableado alostérico en lugar de bloquear la química directamente, los futuros fármacos podrían privar con mayor eficacia a las bacterias de tuberculosis de la flexibilidad metabólica que necesitan para persistir en el cuerpo humano.
Cita: Huang, E.YW., Kwai, B.X.C., Jiao, W. et al. Structural basis of allosteric activation of Mycobacterium tuberculosis isocitrate lyase 2. Commun Biol 9, 560 (2026). https://doi.org/10.1038/s42003-026-09821-6
Palabras clave: metabolismo de la tuberculosis, regulación alostérica, isocitrato liasa 2, señalización por acetil-CoA, blancos farmacológicos antimicrobianos