Clear Sky Science · zh
攀升纤维招募去抑制以增强浦肯野细胞钙信号
大脑如何从错误中学习
小脑是位于大脑后部、拳头大小的结构,帮助我们学习流畅且时机精确的动作,从恰当时刻眨眼到击打网球。本文探讨了一个令人困惑的问题:当教学信号进入小脑时,为什么它们不会被通常抑制脑活动的附近制动信号淹没?通过发现一条短暂解除这一制动的隐藏回路,作者解释了教学信号如何变得足够突出以驱动学习。
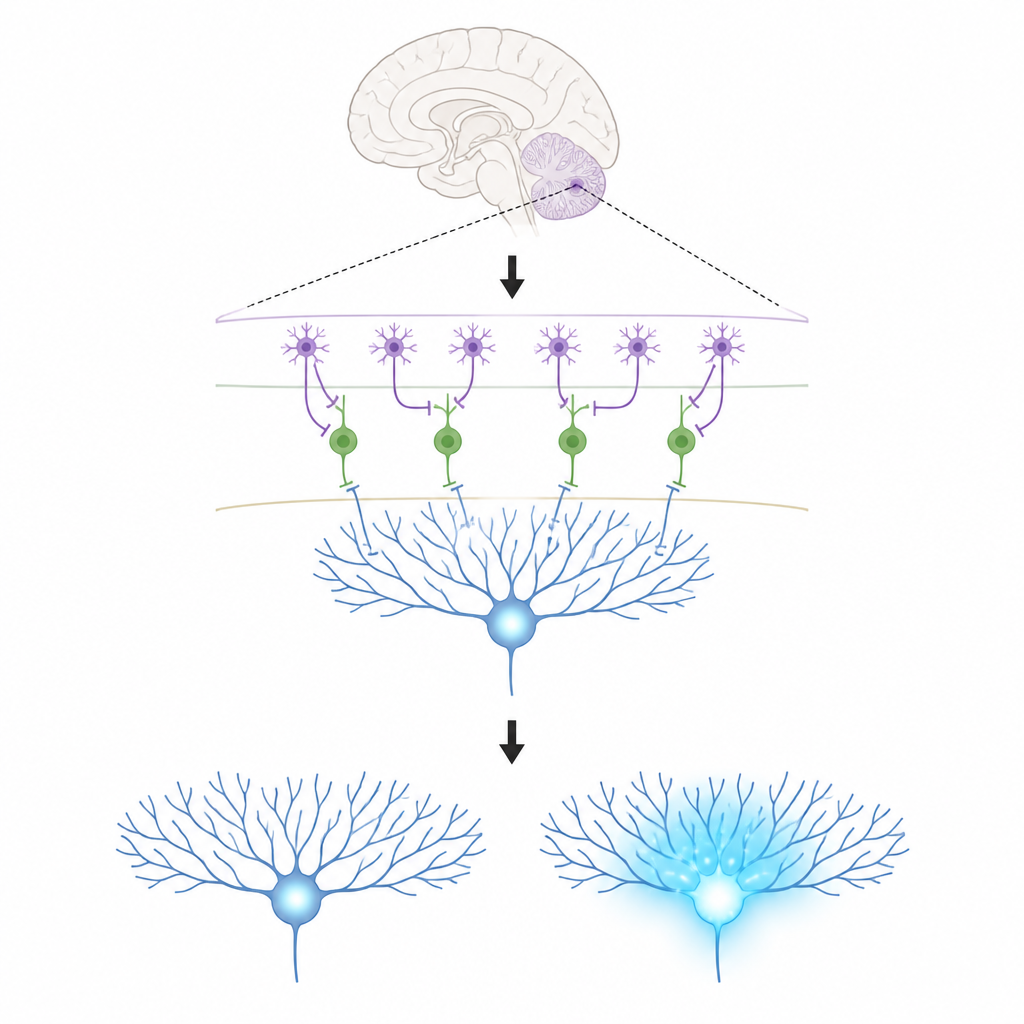
微小回路中的关键参与者
这项工作聚焦于小脑皮层中的三类细胞。浦肯野细胞位于该区域的输出端,帮助塑造我们的动作;它们通过改变所接收突触的强度来学习。攀升纤维携带强大的教学信号,会使浦肯野细胞树突出现大量钙流入,这种化学变化与突触长期调整相关。在薄薄的外层,分子层抑制性中间神经元有两种类型。一类(MLI1)直接抑制浦肯野细胞,起到局部制动的作用。另一类(MLI2)抑制这些制动细胞,有效地释放制动,使浦肯野细胞能够更自由地放电。
教学信号传播中的悖论
已知攀升纤维会强烈激活浦肯野细胞,但早期研究也显示它们会激活那些抑制浦肯野细胞的中间神经元。乍看之下,这似乎自相矛盾:如果同一信号既触发教学脉冲又额外打开制动,浦肯野细胞的钙上升应会被削弱,从而妨碍学习。作者使用详细的电子显微镜重建来追踪小鼠小脑中攀升纤维与两种中间神经元之间的物理接触。他们发现,尽管每条攀升纤维到达两类中间神经元的数量相似,但它在去抑制的MLI2细胞上形成了更多、更大的接触位点,而在直接抑制的MLI1细胞上的接触较少且较小。
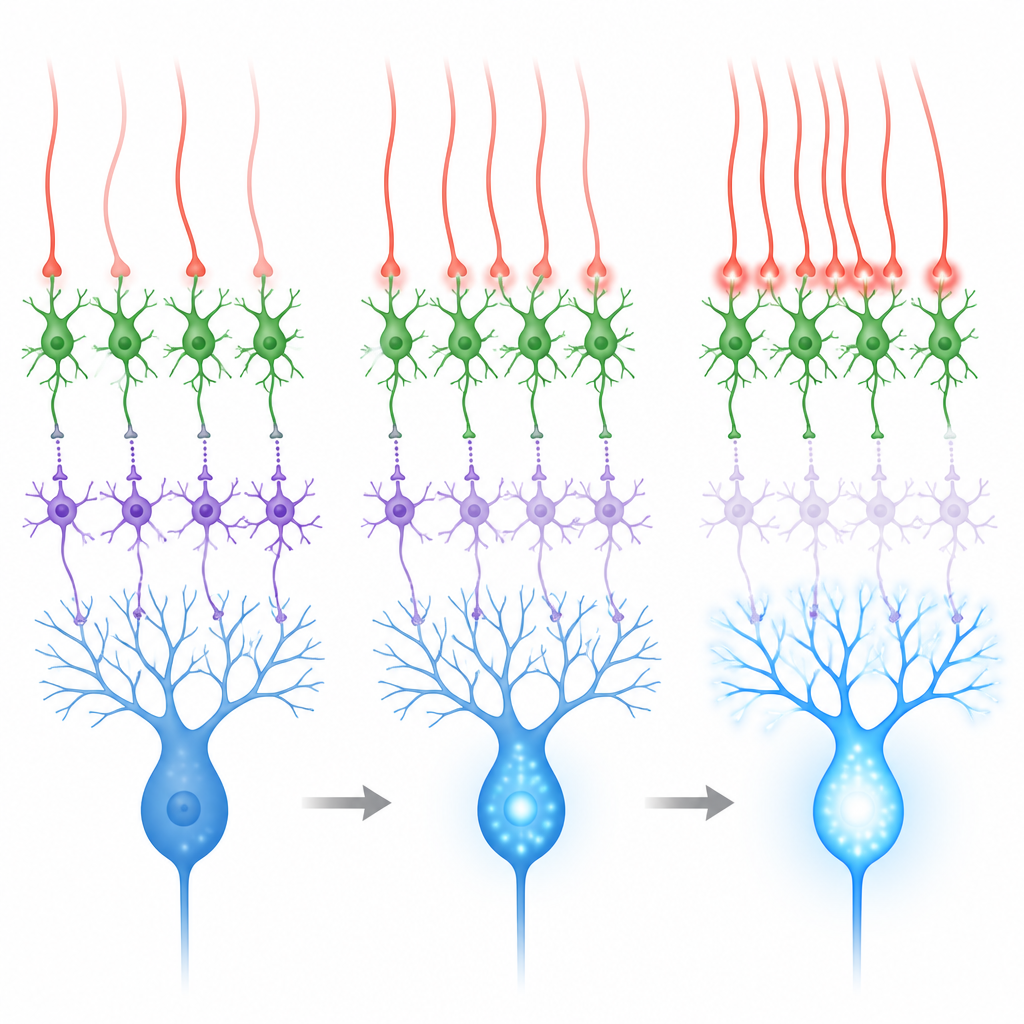
偏向去抑制的溢出信号
这些接触看起来不同寻常:与经典的快速突触不同,它们缺乏在普通突触处常见的紧密囊泡和受体簇。相反,它们位于许多颗粒细胞突触的附近,提示攀升纤维释放的谷氨酸可以扩散或溢出,达到邻近中间神经元上的受体。在脑片实验中,团队在刺激攀升纤维时记录了被鉴定的MLI1和MLI2细胞的电响应。MLI2细胞反应强烈且可靠,表现为大而缓的电流,其中包括对与可塑性相关的一类谷氨酸受体的显著贡献。MLI1细胞的反应较弱,且需要更高的外溶液钙浓度才能清晰观察到。这些数据表明,来自攀升纤维的溢出信号强烈偏向去抑制性的MLI2通路。
从详细电路到行为的联系
为了解在活体大脑中这一机制如何发挥作用,研究者使用高密度Neuropixels探针在清醒奔跑的小鼠中同时记录大量细胞。自发的攀升纤维事件会先在MLI2细胞中产生快速的活动爆发,随后MLI1放电出现延迟下降,浦肯野细胞放电则出现反弹性增加,这与去抑制一致。当团队用受其解剖学和脑片数据约束的计算模型模拟该回路时,观察到相同的模式:攀升纤维输入兴奋MLI2,MLI2抑制许多MLI1,从而在一条窄带组织内解除对附近浦肯野细胞的抑制。
感官信号与同步性的力量
当攀升纤维对有意义的感官事件(例如用于眼睑条件作用任务的眼部气流刺激)发放时,小脑的学习效果最佳。在此类气流刺激期间,教学通路(经攀升纤维)和另一条输入通路(经苔藓纤维和颗粒细胞)都会被激活。第二条通路倾向于同时兴奋两类中间神经元,可能加强对浦肯野细胞的抑制,从而与学习相抗衡。记录和模拟显示,当攀升纤维较为独立地发放时,对浦肯野细胞的净效应是混合的。但当一片相邻的攀升纤维在几毫秒内同步发放时,MLI2细胞被强烈驱动,MLI1细胞被更强地抑制,浦肯野细胞树突中的钙激增更为显著。
这对我们如何学习动作意味着什么
通过结合超微结构解剖、脑片记录、体内记录、计算模型和钙成像,该研究揭示了一条解决原始悖论的去抑制环路。攀升纤维并非简单地同时兴奋与抑制浦肯野细胞;相反,它们的连线优先激活一条在教学信号到来时短暂解除抑制的回路。当许多攀升纤维在感官事件期间同时活跃时,这一回路尤其有效,增强浦肯野细胞树突中的钙信号,并创建一个时间窗口,使编码学会动作的突触发生改变。该机制有助于解释为何攀升纤维的同步活动是推动小脑学习的高效动力。
引用: Santos-Valencia, F., Lackey, E.P., Norton, A. et al. Climbing fibres recruit disinhibition to enhance Purkinje cell calcium signals. Nature 653, 455–464 (2026). https://doi.org/10.1038/s41586-026-10220-4
关键词: 小脑学习, 浦肯野细胞, 攀升纤维, 去抑制, 突触可塑性