Clear Sky Science · de
Kletterfasern rekrutieren Disinhibition, um Purkinje-Zell-Calciumsignale zu verstärken
Wie das Gehirn aus Fehlern lernt
Das Kleinhirn, eine faustgroße Struktur im hinteren Teil des Gehirns, hilft uns, glatte, zeitlich präzise Bewegungen zu erlernen – vom rechtzeitigen Blinzeln bis zum Treffen eines Tennisballs. Diese Studie stellt eine rätselhafte Frage: Wenn Lehrsignale im Kleinhirn eintreffen, warum werden sie nicht von benachbarten Bremssignalen übertönt, die normalerweise die Aktivität dämpfen? Indem die Autoren einen verborgenen Schaltkreis aufdecken, der diese Bremse kurzzeitig aufhebt, erklären sie, wie Lehrsignale stark genug hervortreten können, um Lernen zu bewirken.
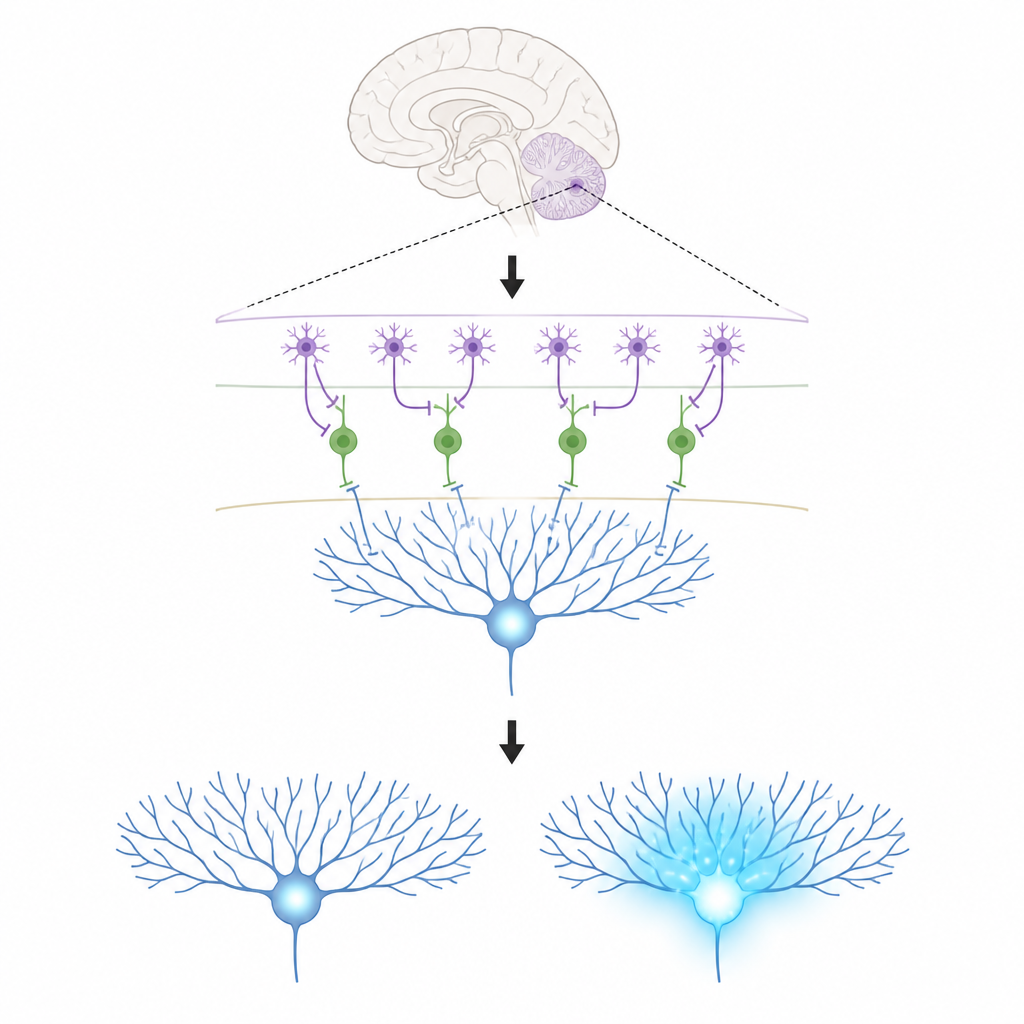
Die Schlüsselrollen in einem winzigen Schaltkreis
Die Arbeit konzentriert sich auf drei Zelltypen in der Kleinhirnrinde. Purkinje-Zellen sitzen am Ausgang dieses Bereichs und formen unsere Bewegungen; sie lernen, indem sie die Stärke der eingehenden Verbindungen ändern. Kletterfasern liefern starke Lehrsignale, die Purkinje-Zellverzweigungen mit Calcium fluten – eine chemische Veränderung, die mit dauerhaften Anpassungen dieser Verbindungen verknüpft ist. In der dünnen äußeren Schicht gibt es zwei Typen molekularer Interneurone. Ein Typ (MLI1) hemmt direkt Purkinje-Zellen und wirkt wie eine lokale Bremse. Der andere Typ (MLI2) hemmt jene Bremszellen, hebt damit die Bremse auf und erlaubt Purkinje-Zellen, freier zu feuern.
Ein Paradoxon in der Ausbreitung von Lehrsignalen
Kletterfasern sind dafür bekannt, Purkinje-Zellen stark zu erregen, doch frühere Arbeiten zeigten auch, dass sie Interneurone erregen, die Purkinje-Zellen hemmen. Auf den ersten Blick wirkt das selbstwidersprüchlich: Wenn dasselbe Signal sowohl einen Lehrimpuls als auch eine zusätzliche Bremse einschaltet, sollte der Calciumanstieg in Purkinje-Zellen kleiner ausfallen und Lernen erschwert werden. Die Autoren nutzten detaillierte Elektronenmikroskop-Rekonstruktionen, um die physischen Kontakte zwischen Kletterfasern und den beiden Interneuron-Typen im Maus-Kleinhirn nachzuzeichnen. Sie fanden, dass, obwohl jede Kletterfaser ähnliche Anzahlen beider Interneuron-Typen erreicht, sie mehr und größere Kontaktstellen zu den disinhibitorischen MLI2-Zellen bildet als zu den direkt hemmenden MLI1-Zellen.
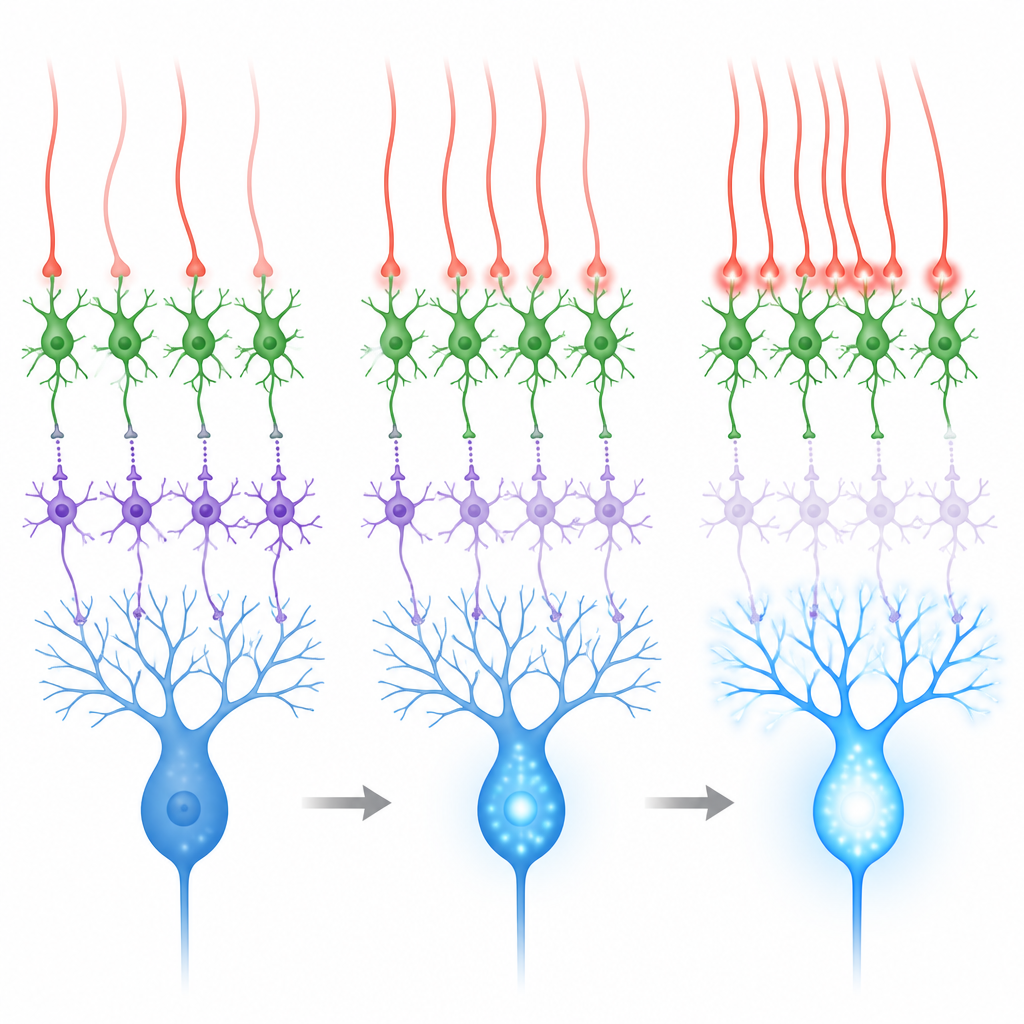
Spillover-Signale, die Disinhibition begünstigen
Diese Kontaktstellen sehen ungewöhnlich aus: Anders als klassische schnelle Synapsen fehlen ihnen die dichten Vesikel- und Rezeptorcluster, die man an gewöhnlichen Verbindungen sieht. Stattdessen liegen sie in der Nähe vieler Körnerzellsynapsen, was darauf hindeutet, dass von Kletterfasern freigesetztes Glutamat diffundieren oder überschwappen kann, um Rezeptoren benachbarter Interneurone zu erreichen. In Schnittpräparaten zeichneten die Forschenden elektrische Antworten identifizierter MLI1- und MLI2-Zellen auf, während sie Kletterfasern stimulierten. MLI2-Zellen reagierten stark und zuverlässig mit großen, langsamen Strömen, zu denen ein wesentlicher Beitrag einer Glutamatrezeptorklasse gehört, die mit Plastizität verknüpft ist. MLI1-Zellen zeigten schwächere Reaktionen und waren nur bei höherem Kalziumgehalt im Bad klar erkennbar. Diese Daten zeigen, dass das Spillover von Kletterfasern eindeutig den disinhibitorischen MLI2-Weg bevorzugt.
Von detaillierter Verschaltung zum Verhalten
Um zu prüfen, wie sich das im lebenden Gehirn auswirkt, setzten die Forschenden dichte Neuropixels-Sonden ein, um viele Zellen gleichzeitig in wachen, laufenden Mäusen aufzuzeichnen. Spontane Kletterfaser-Ereignisse erzeugten einen schnellen Aktivitätsausbruch in MLI2-Zellen, gefolgt von einem verzögerten Abfall der MLI1-Feuerung und einem anschließenden Anstieg der Purkinje-Zell-Spiking-Rate – konsistent mit Disinhibition. Als das Team den Schaltkreis in einem Computer-Modell nachbildete, das durch ihre anatomischen und Schnittdaten eingeschränkt war, beobachteten sie dasselbe Muster: Kletterfaser-Eingang erregt MLI2s, die viele MLI1s ausschalten und damit die Hemmung benachbarter Purkinje-Zellen in einem engen Gewebeband aufheben.
Sensorische Signale und die Kraft der Synchronität
Das Kleinhirn lernt am besten, wenn Kletterfasern als Reaktion auf bedeutungsvolle sensorische Ereignisse feuern, etwa einen Luftstoß aufs Auge, wie er in Lidkonditionierungsaufgaben verwendet wird. Während solcher Luftstöße sind sowohl der Lehrpfad (via Kletterfasern) als auch ein separater Eingangsweg (via Moosfasern und Körnerzellen) aktiv. Der zweite Pfad tendiert dazu, beide Interneuron-Typen zu erregen und kann die Hemmung von Purkinje-Zellen verstärken, was dem Lernen entgegenwirkt. Aufzeichnungen und Simulationen zeigten, dass bei weitgehend unabhängigen Kletterfaser-Feuerungen der Nettoeffekt auf Purkinje-Zellen gemischt ist. Wenn jedoch Gruppen benachbarter Kletterfasern innerhalb weniger Millisekunden gemeinsam feuern, werden MLI2-Zellen stark angetrieben, MLI1-Zellen stärker unterdrückt und Purkinje-Zellen zeigen größere Calciumspitzen in ihren Dendriten.
Was das für das Erlernen von Bewegungen bedeutet
Durch die Kombination von ultrastruktureller Anatomie, Schnittpräparat-Aufzeichnungen, in vivo-Messungen, Computermodellen und Calcium-Imaging deckt die Studie eine disinhibitorische Rückkopplungsschleife auf, die das ursprüngliche Paradoxon löst. Kletterfasern erregen und hemmen Purkinje-Zellen nicht einfach gleichzeitig; stattdessen sind sie so verschaltet, dass sie bevorzugt einen Schaltkreis aktivieren, der die Hemmung genau dann kurz entfernt, wenn ein Lehrsignal eintrifft. Wenn viele Kletterfasern während eines sensorischen Ereignisses zusammen aktiv sind, ist dieser Schaltkreis besonders wirksam, verstärkt Calciumsignale in Purkinje-Dendriten und schafft ein Zeitfenster, in dem die Synapsen, die gelernte Bewegungen kodieren, verändert werden können. Dieser Mechanismus hilft zu erklären, warum synchronisierte Kletterfaser-Aktivität ein so effizienter Treiber des Kleinhirnlernens ist.
Zitation: Santos-Valencia, F., Lackey, E.P., Norton, A. et al. Climbing fibres recruit disinhibition to enhance Purkinje cell calcium signals. Nature 653, 455–464 (2026). https://doi.org/10.1038/s41586-026-10220-4
Schlüsselwörter: Kleinhirnlernen, Purkinje-Zellen, Kletterfasern, Disinhibition, synaptische Plastizität