Clear Sky Science · ru
Восходящие волокна привлекают дисинhibition для усиления кальциевых сигналов нейронов Пуркинье
Как мозг учится на ошибках
Мозжечок, структура размером с кулак в задней части мозга, помогает нам вырабатывать гладкие, точно скоординированные движения — от моргания в нужный момент до удара по теннисному мячу. В этой работе поставлен любопытный вопрос: когда в мозжечок поступают сигналы-«преподавания», почему они не теряются среди соседних тормозящих сигналов, которые обычно подавляют активность? Описав скрытую цепь, которая кратковременно снимает этот тормоз, авторы объясняют, как сигналы обучения могут выделяться достаточно сильно, чтобы вызывать обучение.
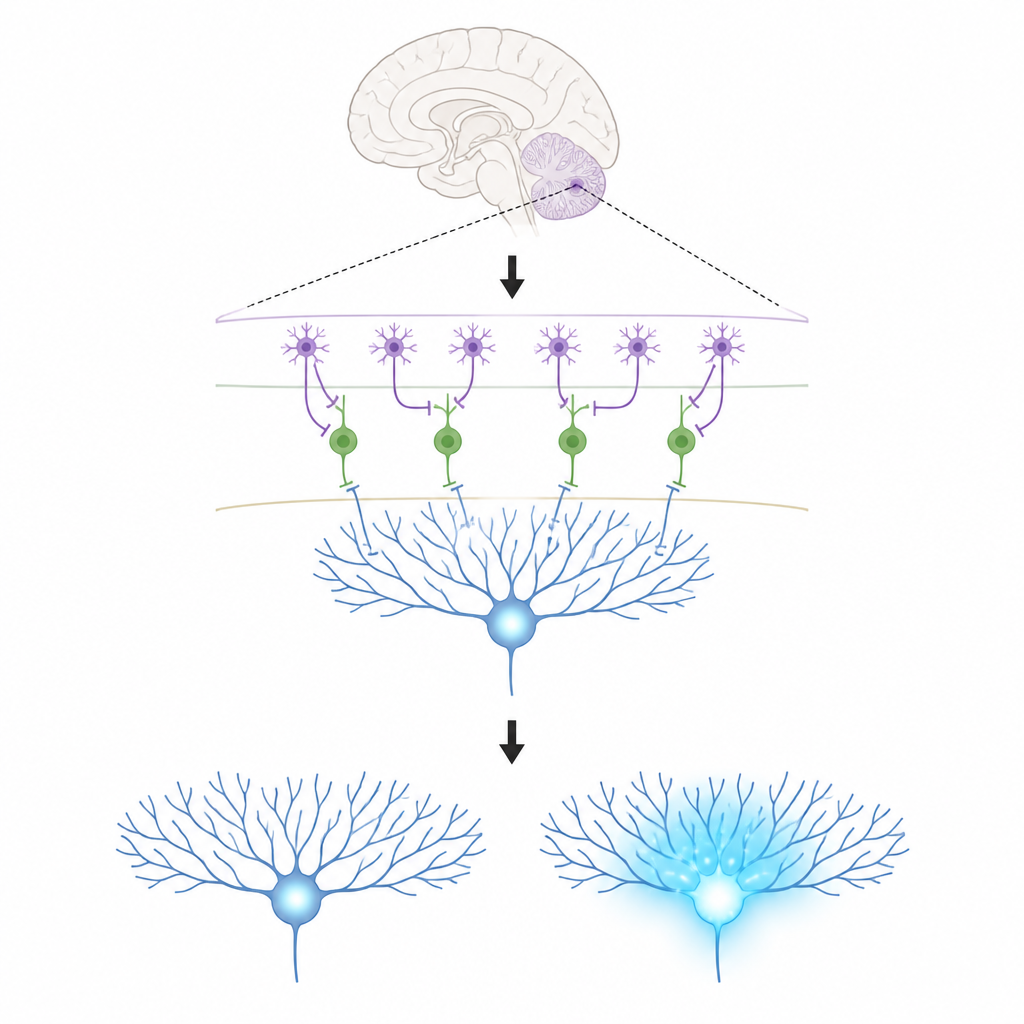
Ключевые участники в крошечной схемe мозга
Работа сосредоточена на трех типах клеток в коре мозжечка. Нейроны Пуркинье находятся на выходе этой области и формируют наши движения; они учатся, изменяя силу получаемых соединений. Восходящие волокна приносят мощные сигналы обучения, заливающие ветви нейронов Пуркинье кальцием — химическим событием, связанным с долговременными изменениями этих связей. В тонком наружном слое молекулярной ткани находятся интернейроны двух типов. Один тип (MLI1) непосредственно тормозит нейроны Пуркинье, выступая как местный «тормоз». Другой тип (MLI2) тормозит эти тормозящие клетки, фактически снимая тормоз и позволяя нейронам Пуркинье активироваться свободнее.
Парадокс в распространении сигнала обучения
Известно, что восходящие волокна сильно возбуждают нейроны Пуркинье, но ранние исследования также показали, что они возбуждают интернейроны, которые тормозят Пуркинье. На первый взгляд это саморазрушающе: если один и тот же сигнал включает одновременно импульс обучения и дополнительный тормоз, повышение кальция в Пуркинье должно уменьшиться, затрудняя обучение. Авторы использовали детальные реконструкции на уровне электронного микроскопа, чтобы проследить физические контакты между восходящими волокнами и двумя типами интернейронов в мозжечке мыши. Они обнаружили, что хотя каждое восходящее волокно достигает похожего числа обоих типов интернейронов, оно образует больше и более крупные контактные площадки на дисинхибиторных клетках MLI2, чем на напрямую тормозящих MLI1.
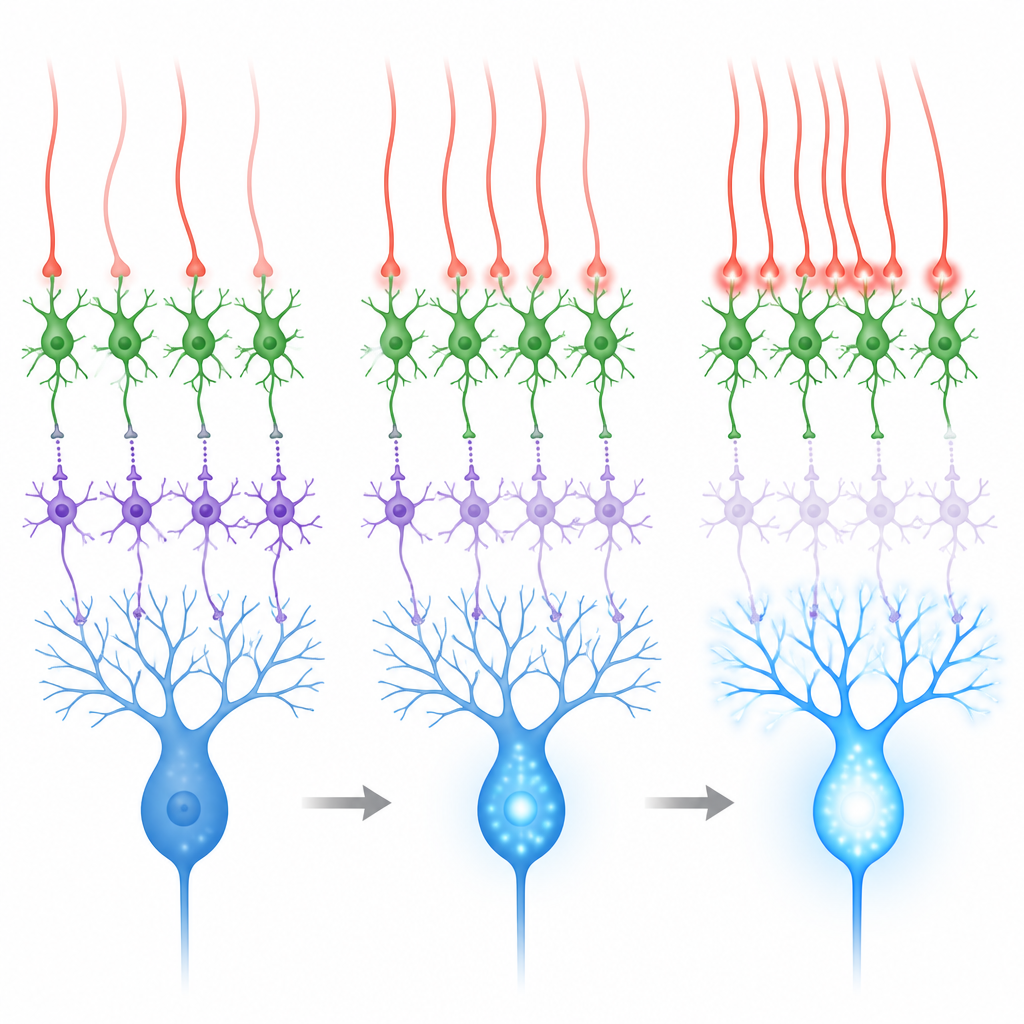
«Просачивающиеся» сигналы в пользу дисинхибиции
Эти контакты выглядят необычно: в отличие от классических быстрых синапсов, им свойственна нехватка плотных скоплений везикул и рецепторных структур, характерных для обычных соединений. Вместо этого они расположены близко к множеству синапсов гранулярных клеток, что предполагает, что глутамат, выделяемый восходящими волокнами, может диффундировать, или «просачиваться», достигая рецепторов на соседних интернейронах. В нервных срезах команда записывала электрические ответы идентифицированных MLI1 и MLI2 при стимуляции восходящих волокон. Клетки MLI2 отвечали сильно и надежно, с большими и медленными токами, в которых была значительная компонента от одного класса глутаматных рецепторов, связанного с пластичностью. MLI1 отвечали слабее и их ответы было легче увидеть при повышенном содержании кальция в растворе. Эти данные показывают, что «просачивание» от восходящих волокон явно благоприятствует пути через дисинхибиторные MLI2.
От детальной схемы к поведению
Чтобы выяснить, как это проявляется в живом мозге, исследователи использовали высокоплотные зонды Neuropixels для одновременной записи многих клеток у бодрствующих бегающих мышей. Спонтанные события восходящих волокон вызывали быстрый взрыв активности в MLI2, за которым следовал задержанный спад в частоте MLI1 и последующее возрастание спайкинга нейронов Пуркинье, что соответствует дисинхибиции. Когда команда смоделировала цепь в компьютере, ограничив модель их анатомическими и срезовыми данными, они получили тот же паттерн: вход восходящих волокон возбуждает MLI2, которые подавляют множество MLI1, снимая тормоз с соседних нейронов Пуркинье в узкой полосе ткани.
Сенсорные сигналы и сила синхронности
Мозжечок лучше всего учится, когда восходящие волокна активируются в ответ на значимые сенсорные события, например на воздушный толчок по глазу, используемый в задачах обусловливания века. Во время таких толчков активируются и путь преподавания (через восходящие волокна), и отдельный входной путь (через моховидные волокна и гранулярные клетки). Второй путь имеет тенденцию возбуждать оба типа интернейронов и может усиливать торможение нейронов Пуркинье, работая против обучения. Записи и симуляции показали, что когда восходящие волокна стреляют более или менее независимо, суммарный эффект на Пуркинье получается смешанным. Но когда группы соседних восходящих волокон активируются вместе в течение нескольких тысячных долей секунды, MLI2 сильно возбуждаются, MLI1 сильнее подавляются, и в ветвях нейронов Пуркинье наблюдаются большие всплески кальция.
Что это значит для того, как мы учим движения
Сочетая ультраструктурную анатомию, записи в срезах, записи in vivo, компьютерные модели и кальциевую визуализацию, исследование раскрывает дисинхибиторную петлю, которая решает исходный парадокс. Восходящие волокна не просто одновременно возбуждают и тормозят нейроны Пуркинье; вместо этого они устроены так, чтобы предпочтительно включать цепь, которая кратковременно снимает ингибирование именно тогда, когда приходит сигнал обучения. Когда много восходящих волокон активны одновременно во время сенсорного события, эта цепь особенно эффективна, усиливая кальциевые сигналы в дендритах Пуркинье и создавая временное окно, в котором синапсы, кодирующие выученные движения, могут изменяться. Этот механизм помогает объяснить, почему синхронизированная активность восходящих волокон так эффективна для обучения в мозжечке.
Цитирование: Santos-Valencia, F., Lackey, E.P., Norton, A. et al. Climbing fibres recruit disinhibition to enhance Purkinje cell calcium signals. Nature 653, 455–464 (2026). https://doi.org/10.1038/s41586-026-10220-4
Ключевые слова: обучение в мозжечке, нейроны Пуркинье, восходящие волокна, дисинхибиция, синаптическая пластичность