Clear Sky Science · ja
登攀線維は脱抑制を動員してプルキンエ細胞のカルシウム信号を増強する
間違いから脳が学ぶ仕組み
小脳は後頭部にあるこぶし大の構造で、瞬きのタイミングからテニスの打球まで、滑らかで適切なタイミングの運動を学習するのに寄与します。本研究は一つの不可解な問いを扱います:教示信号が小脳に到達したとき、通常は活動を抑える近傍のブレーキ信号にかき消されないのはなぜか。著者らはこのブレーキを一時的に解除する隠れた回路を明らかにすることで、教示信号が学習を駆動するのに十分な強さで目立つ仕組みを説明します。
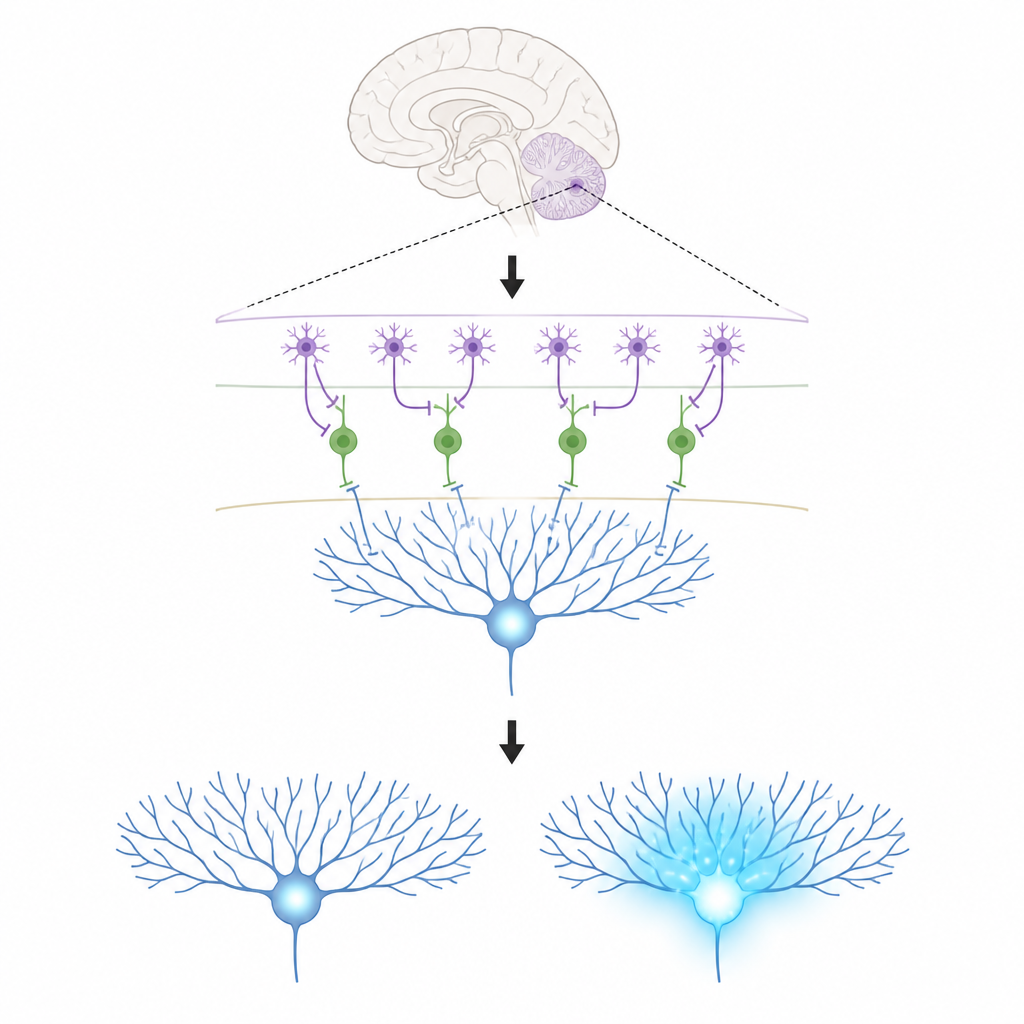
小さな回路の主要な登場人物
この研究は小脳皮質の3種類の細胞に着目します。プルキンエ細胞はこの領域の出力に位置し、運動の形成を助けます;それらは受け取る結合の強さを変えることで学習します。登攀線維は強力な教示信号を運び、プルキンエ細胞の樹状枝にカルシウムを大量に流入させます。カルシウムの変化はそれらの結合に持続的な調整をもたらすことと関連しています。薄い外層では分子層介在ニューロン(MLI)が2種類あり、1つ(MLI1)はプルキンエ細胞を直接抑制して局所のブレーキとして働きます。もう1つ(MLI2)はその抑制細胞を抑えることで実質的にブレーキを解除し、プルキンエ細胞がより自由に発火できるようにします。
教示信号の拡がりにおける逆説
登攀線維はプルキンエ細胞を強力に興奮させることで知られますが、以前の研究はまた登攀線維がプルキンエ細胞を抑制する介在ニューロンも興奮させることを示していました。一見すると自滅的です:同じ信号が教示パルスと追加のブレーキの両方をオンにするなら、プルキンエ細胞のカルシウム上昇は小さくなり、学習が難しくなるはずです。著者らは詳細な電子顕微鏡再構築を用いて、マウス小脳における登攀線維と両介在ニューロン型との物理的接触を追跡しました。各登攀線維は両方の介在ニューロン型にほぼ同数到達しますが、脱抑制を担うMLI2に対しては、直接抑制するMLI1よりもより多く、かつ大きな接触部位を形成していることを見いだしました。
脱抑制を有利にするこぼれ信号(スピルオーバー)
これらの接触はやや異様に見えます:古典的な速いシナプスのような小胞や受容体構造の密なクラスターが欠けています。代わりに多くの顆粒細胞シナプスの近くに位置しており、登攀線維から放出されたグルタミン酸が拡散(スピルオーバー)して近傍の介在ニューロンの受容体に到達する可能性を示唆します。脳スライス内で、研究チームは登攀線維を刺激しつつ同定されたMLI1とMLI2細胞の電気応答を記録しました。MLI2細胞は大きく遅い電流で強く確実に応答し、可塑性に関連する一群のグルタミン酸受容体からの大きな寄与を含んでいました。MLI1細胞の応答はより弱く、明確に観察するには浴液中のカルシウムを高める必要がありました。これらのデータは、登攀線維からのスピルオーバーが脱抑制経路であるMLI2を強く好むことを示しています。
詳細な回路から行動へ
生体内でこれがどのように現れるかを確かめるため、研究者らは高密度Neuropixelsプローブを用いて、覚醒して走行するマウスで多数の細胞を同時に記録しました。自発的な登攀線維イベントはMLI2細胞の速い活動バーストを引き起こし、その後遅れてMLI1発火の低下とプルキンエ細胞のスパイクのリバウンド増加をもたらし、脱抑制と一致しました。解剖学とスライスデータで制約されたコンピュータモデルで回路をシミュレーションすると、同じパターンが観察されました:登攀線維入力はMLI2を興奮させ、多くのMLI1を抑え、それにより狭い組織帯内の近傍プルキンエ細胞から抑制を取り除きます。
感覚信号と同期の力
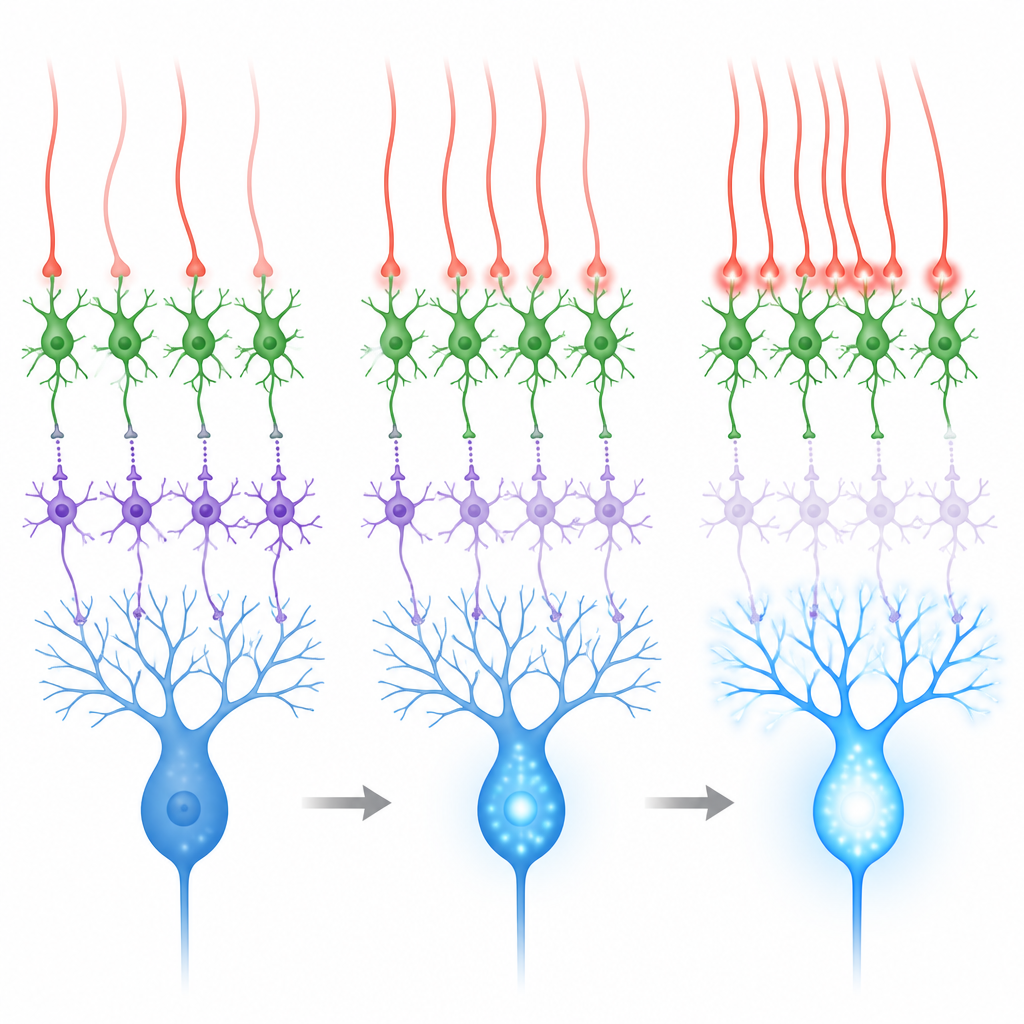
小脳は登攀線維が有意味な感覚事象に応答して発火する場合に最もよく学習します。まぶた条件付け課題で用いられる眼への空気噴射のような事象です。そのような空気噴射では、教示経路(登攀線維経由)と別の入力経路(苔状線維と顆粒細胞経由)の両方が活性化します。後者の経路は両方の介在ニューロン型を興奮させ、プルキンエ細胞の抑制を強化して学習に逆行することがあります。記録とシミュレーションは、登攀線維がばらばらに発火する場合、プルキンエ細胞への純粋な影響は混合的になることを示しました。しかし、近傍の登攀線維群が数ミリ秒以内に同時に発火すると、MLI2細胞が強く駆動され、MLI1細胞はより強く抑制され、プルキンエ細胞の樹状枝でより大きなカルシウム急増が起こります。
運動を学ぶ仕組みへの意味
超微細構造の解剖、スライス記録、生体内記録、計算モデル、カルシウムイメージングを組み合わせることで、本研究は元の逆説を解く脱抑制ループを明らかにします。登攀線維は単にプルキンエ細胞を同時に興奮させたり抑制したりするのではなく、教示信号が到来したちょうどその時に抑制を一時的に解除する回路を優先的にオンにするよう配線されています。感覚事象の際に多数の登攀線維が同時に活性化すると、この回路は特に効果的に働き、プルキンエ細胞樹状枝のカルシウム信号を強化し、学習された運動を符号化するシナプスが変化できる時間窓を生み出します。このメカニズムは、同期した登攀線維活動が小脳学習の効率的な駆動因である理由を説明するのに役立ちます。
引用: Santos-Valencia, F., Lackey, E.P., Norton, A. et al. Climbing fibres recruit disinhibition to enhance Purkinje cell calcium signals. Nature 653, 455–464 (2026). https://doi.org/10.1038/s41586-026-10220-4
キーワード: 小脳学習, プルキンエ細胞, 登攀線維, 脱抑制, シナプス可塑性