Clear Sky Science · pl
Włókna wspinające rekrutują dysinhibicję, by wzmocnić sygnały wapniowe komórek Purkinjego
Jak mózg uczy się na błędach
Móżdżek, struktura wielkości pięści zlokalizowana z tyłu mózgu, pomaga nam opanowywać gładkie, precyzyjnie wyczesane ruchy — od mrugnięcia we właściwym momencie po trafienie w piłkę tenisową. W badaniu postawiono intrygujące pytanie: gdy do móżdżku docierają sygnały uczące, dlaczego nie są zagłuszane przez pobliskie sygnały hamujące, które zwykle tłumią aktywność neuronów? Odkrywając ukryty obwód, który chwilowo podnosi ten hamulec, autorzy wyjaśniają, jak sygnały uczące mogą się wyróżnić na tyle, by napędzać proces uczenia się.
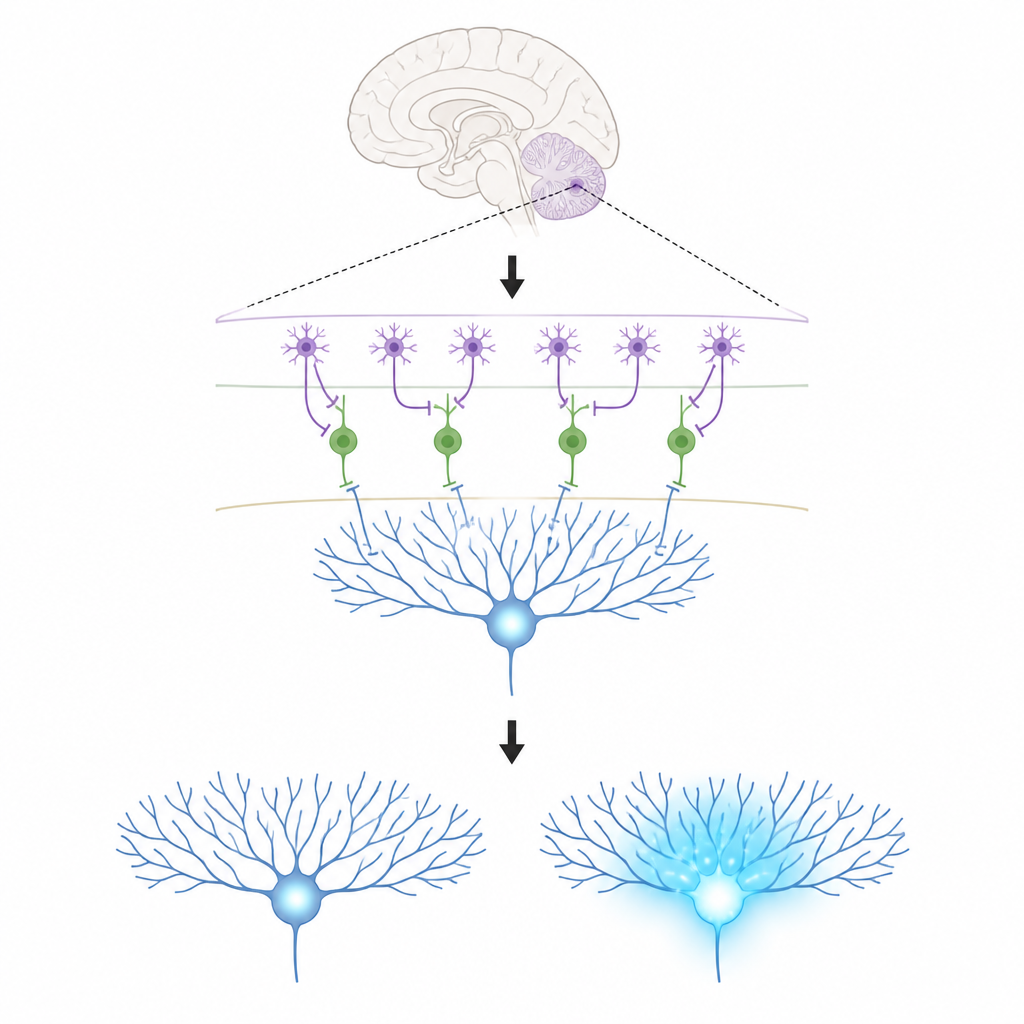
Główni aktorzy w małym obwodzie mózgowym
Prace koncentrują się na trzech typach komórek w korze móżdżku. Komórki Purkinjego znajdują się na wyjściu tego obszaru i kształtują nasze ruchy; uczą się poprzez zmianę siły otrzymywanych połączeń. Włókna wspinające przynoszą silne sygnały uczące, które zalewają rozgałęzienia komórek Purkinjego jonami wapnia — zmiana chemiczna powiązana z trwałymi modyfikacjami tych połączeń. W cienkiej warstwie zewnętrznej znajdują się interneurony warstwy molekularnej w dwóch odmianach. Jeden typ (MLI1) bezpośrednio hamuje komórki Purkinjego, działając jak lokalny hamulec. Drugi typ (MLI2) hamuje te hamujące komórki, skutecznie zwalniając hamulec i pozwalając komórkom Purkinjego wyładowywać się swobodniej.
Pozór paradoksu w rozprzestrzenianiu się sygnału uczącego
Wiadomo, że włókna wspinające silnie pobudzają komórki Purkinjego, lecz wcześniejsze badania pokazywały też, że pobudzają interneurony hamujące komórki Purkinjego. Na pierwszy rzut oka wydaje się to samoniszczące: jeśli ten sam sygnał uruchamia zarówno impuls uczący, jak i dodatkowy hamulec, wzrost wapnia w komórkach Purkinjego powinien słabnąć, utrudniając uczenie się. Autorzy wykorzystali szczegółowe rekonstrukcje w mikroskopii elektronowej, aby prześledzić fizyczne kontakty między włóknami wspinającymi a oboma typami interneuronów w móżdżku myszy. Stwierdzili, że chociaż każde włókno wspinające sięga podobnej liczby obu typów interneuronów, tworzy więcej i większych miejsc kontaktu z dysinhibicyjnymi komórkami MLI2 niż z bezpośrednio hamującymi MLI1.
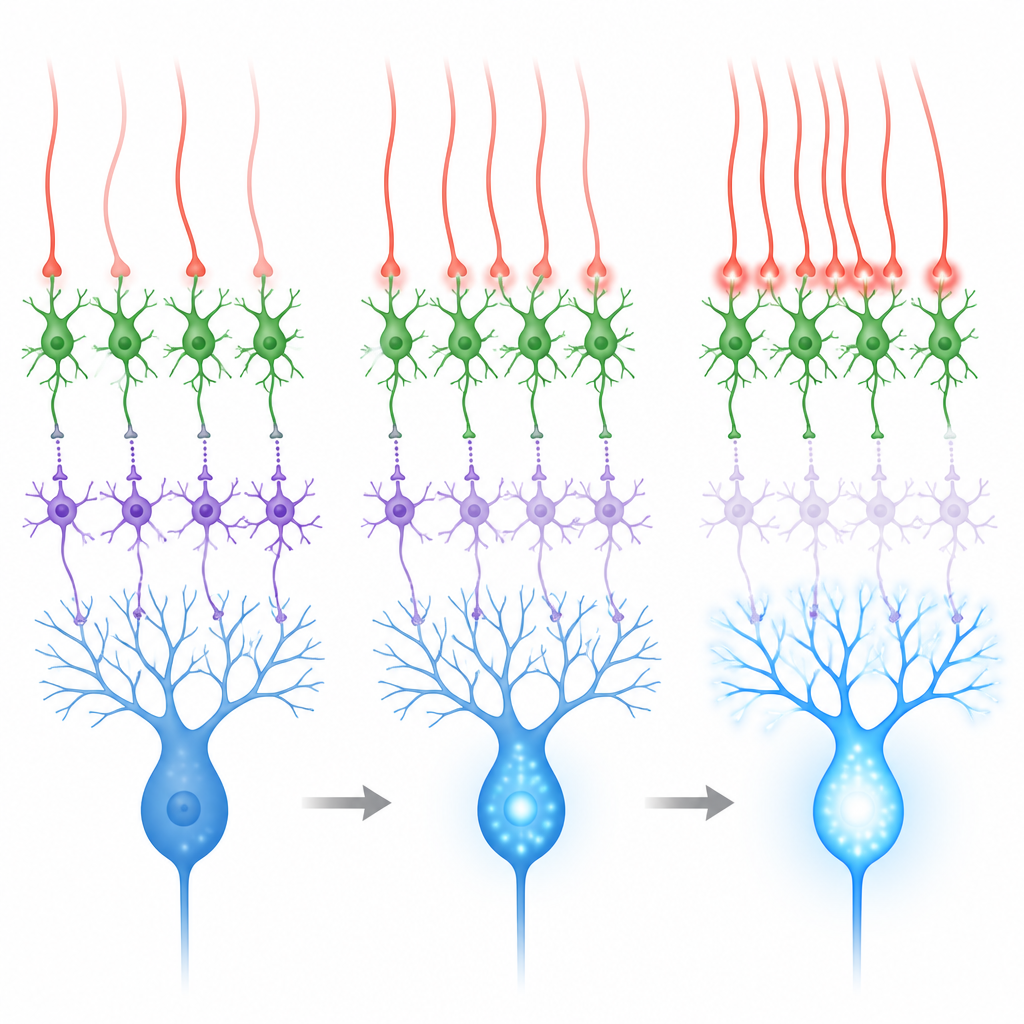
Przelew sygnałów sprzyjający dysinhibicji
Te miejsca kontaktu wyglądają nietypowo: w odróżnieniu od klasycznych szybkich synaps, brakuje im ciasnych skupisk pęcherzyków i struktur receptorowych widocznych w zwykłych złączach. Zamiast tego leżą blisko wielu synaps komórek ziarnistych, co sugeruje, że glutaminian uwalniany z włókien wspinających może dyfundować, czyli „przelewać się”, sięgając receptorów na pobliskich interneuronach. W preparatach skrawków mózgu zespół rejestrował odpowiedzi elektryczne zidentyfikowanych komórek MLI1 i MLI2 podczas stymulacji włókien wspinających. Komórki MLI2 odpowiadały silnie i niezawodnie, dużymi, wolnymi prądami zawierającymi istotny wkład jednego typu receptorów glutaminianu powiązanych z plastycznością. Komórki MLI1 reagowały słabiej i ich odpowiedzi były widoczne wyraźniej dopiero przy wyższym stężeniu wapnia w roztworze. Dane te pokazują, że przelew z włókien wspinających silnie preferuje ścieżkę dysinhibicyjną MLI2.
Od szczegółowej anatomii obwodu do zachowania
Aby sprawdzić, jak to działa w żywym mózgu, badacze użyli sond Neuropixels o wysokiej gęstości, by rejestrować wiele komórek jednocześnie u czujnych, biegających myszy. Spontaniczne zdarzenia włókien wspinających wywoływały szybki wybuch aktywności w komórkach MLI2, po którym następował opóźniony spadek wyładowań MLI1 i odbiciowy wzrost wyładowań komórek Purkinjego — zgodny z mechanizmem dysinhibicji. Gdy zespół zasymulował obwód w modelu komputerowym ograniczonym przez ich dane anatomiczne i skrawkowe, zaobserwowano ten sam wzorzec: wejście włókien wspinających pobudza MLI2, które wyłączają wiele MLI1, podnosząc hamowanie z pobliskich komórek Purkinjego w wąskim pasie tkanki.
Sygnały sensoryczne i siła synchronii
Móżdżek najlepiej się uczy, gdy włókna wspinające wyładowują się w odpowiedzi na istotne zdarzenia sensoryczne, na przykład pchnięcie powietrza w oko wykorzystywane w zadaniach kondycjonowania powiek. Podczas takich bodźców aktywne są zarówno szlak uczący (przez włókna wspinające), jak i odrębna droga wejścia (przez włókna mszyste i komórki ziarniste). Drugi szlak ma tendencję do pobudzania obu typów interneuronów i może wzmacniać hamowanie komórek Purkinjego, działając przeciw uczącemu sygnałowi. Nagrania i symulacje wykazały, że gdy włókna wspinające aktywują się mniej więcej niezależnie, efekt netto na komórki Purkinjego jest mieszany. Gdy jednak grupy pobliskich włókien wspinających wystrzeliwują razem w ciągu kilku tysięcznych sekundy, komórki MLI2 są silnie napędzane, MLI1 są mocniej tłumione, a komórki Purkinjego wykazują większe skoki wapniowe w swoich gałęziach.
Co to oznacza dla uczenia się ruchów
Łącząc ultrastrukturalną anatomię, nagrania ze skrawków, nagrania in vivo, modele komputerowe i obrazowanie wapniowe, badanie ujawnia pętlę dysinhibicyjną, która rozwiązuje pierwotny paradoks. Włókna wspinające nie tylko jednocześnie pobudzają i hamują komórki Purkinjego; są połączone tak, by uprzywilejować włączanie obwodu, który chwilowo usuwa hamowanie dokładnie wtedy, gdy przychodzi sygnał uczący. Gdy wiele włókien wspinających działa synchronicznie podczas zdarzenia sensorycznego, mechanizm ten jest szczególnie skuteczny, wzmacniając sygnały wapniowe w dendrytach komórek Purkinjego i tworząc okno czasowe, w którym synapsy kodujące wyuczone ruchy mogą ulec zmianie. Ten mechanizm pomaga wyjaśnić, dlaczego zsynchronizowana aktywność włókien wspinających jest tak efektywnym motorem uczenia móżdżkowego.
Cytowanie: Santos-Valencia, F., Lackey, E.P., Norton, A. et al. Climbing fibres recruit disinhibition to enhance Purkinje cell calcium signals. Nature 653, 455–464 (2026). https://doi.org/10.1038/s41586-026-10220-4
Słowa kluczowe: uczenie móżdżkowe, komórki Purkinjego, włókna wspinające, dysinhibicja, plastyczność synaptyczna