Clear Sky Science · en
Climbing fibres recruit disinhibition to enhance Purkinje cell calcium signals
How the Brain Learns from Mistakes
The cerebellum, a fist sized structure at the back of the brain, helps us learn smooth, well timed movements, from blinking at the right moment to hitting a tennis ball. This study asks a puzzling question: when teaching signals arrive in the cerebellum, why do they not get drowned out by nearby braking signals that normally quiet brain activity? By uncovering a hidden circuit that briefly lifts this brake, the authors explain how teaching signals can stand out strongly enough to drive learning.
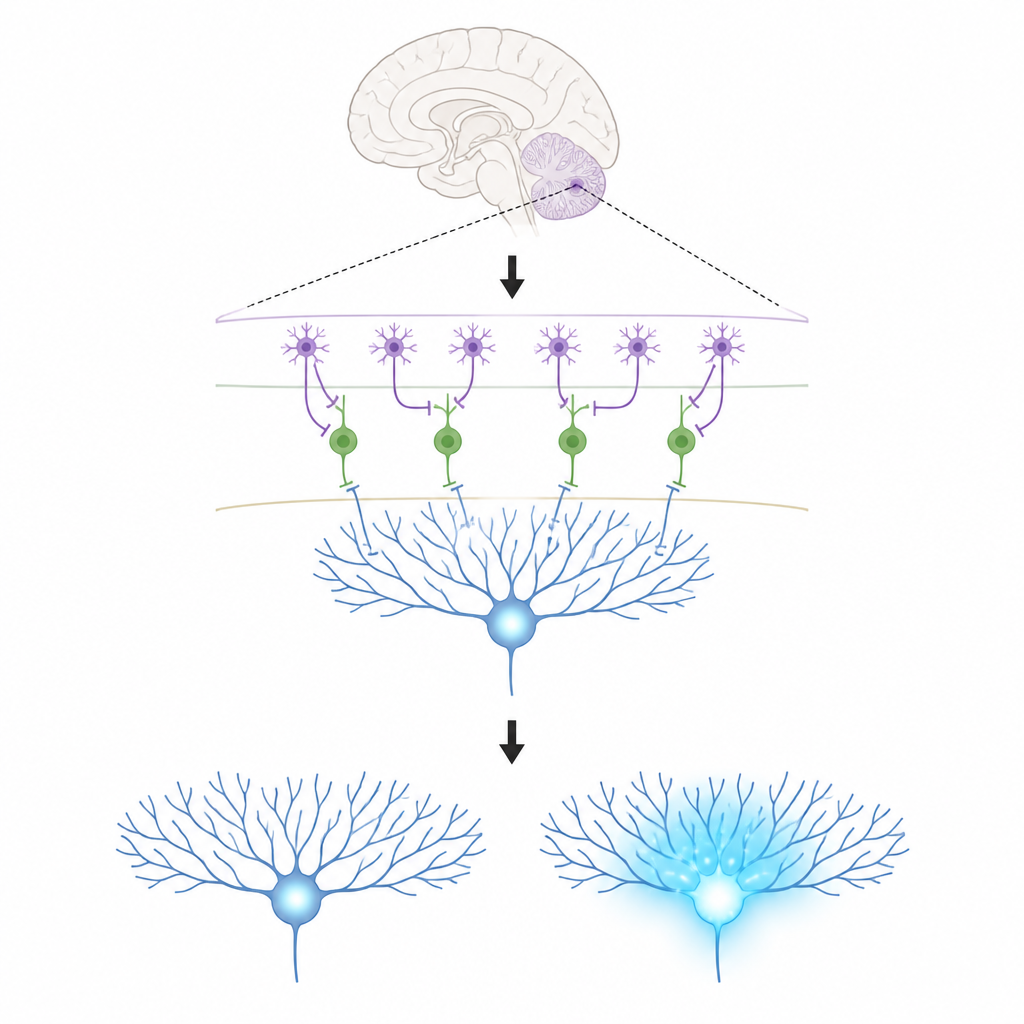
The Key Players in a Tiny Brain Circuit
The work focuses on three kinds of cells in the cerebellar cortex. Purkinje cells sit at the output of this region and help shape our movements; they learn by changing the strength of the connections they receive. Climbing fibres bring powerful teaching signals that flood Purkinje cell branches with calcium, a chemical change linked to lasting adjustments in those connections. In the thin outer layer, molecular layer interneurons come in two types. One type (MLI1) directly inhibits Purkinje cells, acting like a local brake. The other type (MLI2) inhibits those braking cells, effectively releasing the brake and allowing Purkinje cells to fire more freely.
A Paradox in How Teaching Signals Spread
Climbing fibres are known to powerfully excite Purkinje cells, but earlier work also showed that they excite interneurons that inhibit Purkinje cells. At first glance, this is self defeating: if the same signal turns on both a teaching pulse and an extra brake, the calcium rise in Purkinje cells should shrink, making learning harder. The authors used detailed electron microscope reconstructions to trace the physical contacts between climbing fibres and the two interneuron types in mouse cerebellum. They found that while each climbing fibre reaches similar numbers of both interneuron types, it forms more, larger contact sites onto the disinhibiting MLI2 cells than onto the directly inhibiting MLI1 cells.
Spillover Signals that Favor Disinhibition
Those contacts look unusual: unlike classic fast synapses, they lack the tight clusters of vesicles and receptor structures seen at ordinary junctions. Instead, they sit close to many granule cell synapses, suggesting that glutamate released from climbing fibres can diffuse, or spill over, to reach receptors on nearby interneurons. In brain slices, the team recorded electrical responses from identified MLI1 and MLI2 cells while stimulating climbing fibres. MLI2 cells responded strongly and reliably, with large and slow currents that included a big contribution from one class of glutamate receptor linked to plasticity. MLI1 cells responded more weakly and required higher calcium in the bathing solution to be seen clearly. These data show that spillover from climbing fibres strongly prefers the disinhibiting MLI2 route.
From Detailed Circuitry to Behavior
To see how this plays out in the living brain, the researchers used high density Neuropixels probes to record many cells at once in awake, running mice. Spontaneous climbing fibre events produced a quick burst of activity in MLI2 cells, followed by a delayed drop in MLI1 firing and a rebound increase in Purkinje cell spiking, consistent with disinhibition. When the team simulated the circuit using a computer model constrained by their anatomy and slice data, they observed the same pattern: climbing fibre input excites MLI2s, which shut down many MLI1s, lifting inhibition from nearby Purkinje cells within a narrow band of tissue.
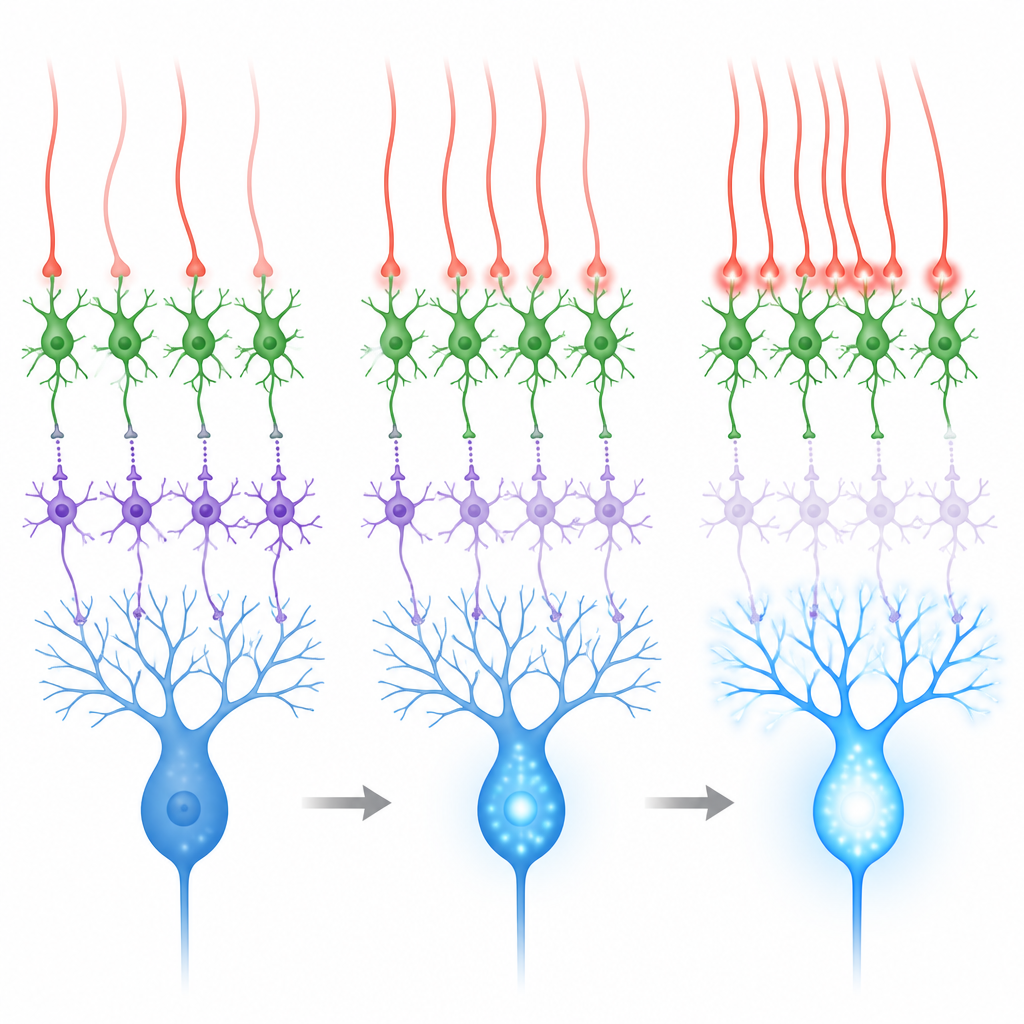
Sensory Signals and the Power of Synchrony
The cerebellum learns best when climbing fibres fire in response to meaningful sensory events, such as an air puff to the eye used in eyelid conditioning tasks. During such air puffs, both the teaching pathway (via climbing fibres) and a separate input pathway (via mossy fibres and granule cells) become active. The second pathway tends to excite both interneuron types and can reinforce inhibition of Purkinje cells, working against learning. Recordings and simulations showed that when climbing fibres fire more or less independently, the net effect on Purkinje cells is mixed. But when groups of nearby climbing fibres fire together within a few thousandths of a second, MLI2 cells are strongly driven, MLI1 cells are more strongly suppressed, and Purkinje cells show larger calcium surges in their branches.
What This Means for How We Learn Movements
By combining ultrastructural anatomy, slice recordings, in vivo recordings, computer models and calcium imaging, the study reveals a disinhibitory loop that solves the original paradox. Climbing fibres do not simply excite and inhibit Purkinje cells at the same time; instead, they are wired to preferentially turn on a circuit that briefly removes inhibition just when a teaching signal arrives. When many climbing fibres are active together during a sensory event, this circuit is especially effective, boosting calcium signals in Purkinje cell dendrites and creating a time window in which the synapses that encode learned movements can change. This mechanism helps explain why synchronized climbing fibre activity is such an efficient driver of cerebellar learning.
Citation: Santos-Valencia, F., Lackey, E.P., Norton, A. et al. Climbing fibres recruit disinhibition to enhance Purkinje cell calcium signals. Nature 653, 455–464 (2026). https://doi.org/10.1038/s41586-026-10220-4
Keywords: cerebellar learning, Purkinje cells, climbing fibers, disinhibition, synaptic plasticity