Clear Sky Science · ru
Молекулярный и функциональный разбор с помощью CaMPARI-seq выявляет организацию нейронов, отвечающих за различение зависимых от оптического потока поведенческих реакций
Как мозг отличает поворот от прямого плавания
Когда животное движется, весь визуальный мир кажется текущим мимо его глаз. Из этой постоянно меняющейся сцены мозг должен извлечь простой, но жизненно важный вопрос: мы поворачиваемся или движемся прямо? В этой работе на крошечных зебрафишах и с помощью новой молекулярной «подсветки» авторы показывают, как конкретные нейроны разделяют эти два вида движения, чтобы управлять либо движениями глаз, либо движениями всего тела.
Восприятие движения как текущего ландшафта
Когда животное плавает или идет, узоры света прокатываются по глазам — это явление называют оптическим потоком. Ротационный поток, например при повороте головы, в основном должен вызывать движения глаз для стабилизации взгляда. Трансляционный поток, как при движении вперед, должен, наоборот, вызывать моторные реакции тела, чтобы двигаться вместе с окружающей водой или поверхностью. У многих позвоночных, включая зебрафиш, центральную роль в интерпретации этих закономерностей движения и направлении сигналов к моторным центрам играет средний мозг — претектум. Предыдущие исследования показали множество претектальных нейронов, реагирующих на оптический поток, но оставалось неясным, какие именно типы клеток, определяемые их экспрессией генов и связями, отвечают за разные поведенческие реакции, вызванные движением.
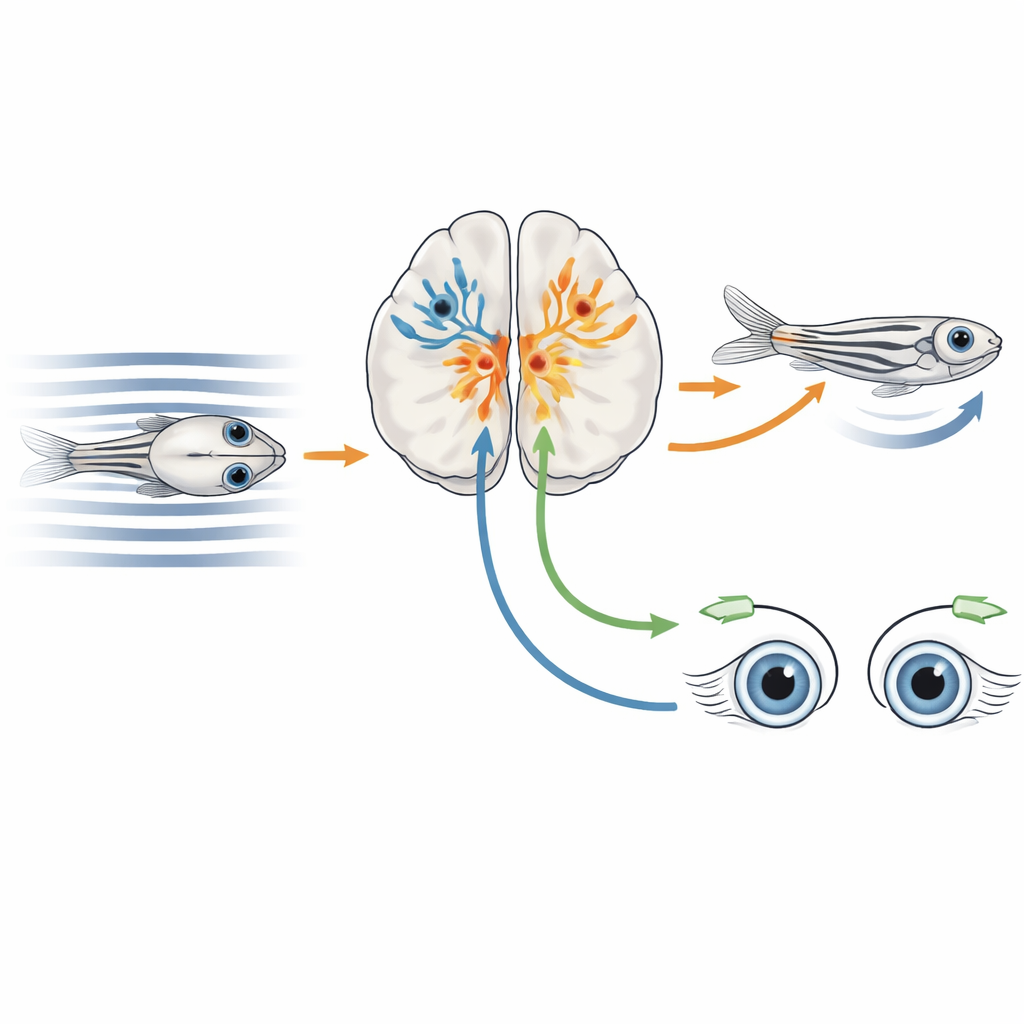
Подсветить активные нейроны и прочитать их гены
Исследователи разработали гибридный подход, который они назвали CaMPARI-seq, сочетая оптическую маркировку активных нейронов с профилированием экспрессии генов в одиночных клетках. Они генетически модифицировали личинок зебрафиш так, чтобы большинство нейронов синтезировали специальный флуоресцентный белок CaMPARI2, нацеленный в ядро клетки. Когда нейроны активны и их подсвечивают ультрафиолетом, этот белок необратимо переключается из зеленого в красный. Команда показывала рыбкам движущиеся полосы, создававшие разнообразные бинокулярные условия оптического потока, и одновременно подсвечивала претектум ультрафиолетом. Активные, реагировавшие на движение нейроны окрашивались в красный, затем их по одной изолировали и секвенировали их РНК, чтобы определить, какие гены экспрессирует каждая клетка.
Построение клеточного атласа нейронов, реагирующих на оптический поток
Кластеризовав тысячи помеченных клеток по шаблонам экспрессии генов, авторы выявили крупную группу в претектуме, маркированную геном tcf7l2, большая часть клеток которой также несла гены, типичные для ингибирующих нейронов. Используя генетически модифицированных рыб, в которых клетки, положительные по tcf7l2, выражали кальциевый индикатор, они проверили, что эта широкая популяция включала почти все ранее описанные типы ответов на оптический поток — от нейронов, настроенных на движение, регистрируемое только одним глазом, до тех, которые реагировали только тогда, когда оба глаза воспринимали согласованное движение вперед или назад. Дальнейшее разделение группы tcf7l2 выявило семь молекулярно отличных субтипов, каждый с собственной комбинацией маркерных генов и преимущественно неперекрывающимся пространственным положением в претектуме, что указывает на мозаичную организацию специализированных ингибирующих схем вокруг узла обработки движения.
Два ключевых субтипа с очень разными задачами
Среди этих субтипов выделялись два. Клетки, экспрессирующие ген mafaa, располагались в латерализованной мигрировавшей зоне, совпадающей с приходящими направленно-селективными волокнами сетчатки. Визуализация и анатомическое трассирование показали, что эти нейроны имеют локальные связи и сильно реагируют, когда полосы движутся в определенном направлении только в одном глазу, что соответствует роли в кодировании простого, глаз-специфического движения, поддерживающего повороты глаз. Напротив, нейроны, экспрессирующие nkx1.2lb, занимали более медиальную зону и отсылали длинные перекрещивающиеся проекции через дорсальную мозговую комиссуру на противоположную сторону, образуя мост между левыми и правыми претектальными цепями. Эти nkx1.2lb-положительные клетки также в основном были ингибирующими и в совокупности покрывали широкий спектр типов ответов на оптический поток, включая как монокулярные, так и бинокулярные паттерны, что указывает на их роль в сравнении сигналов от двух глаз.
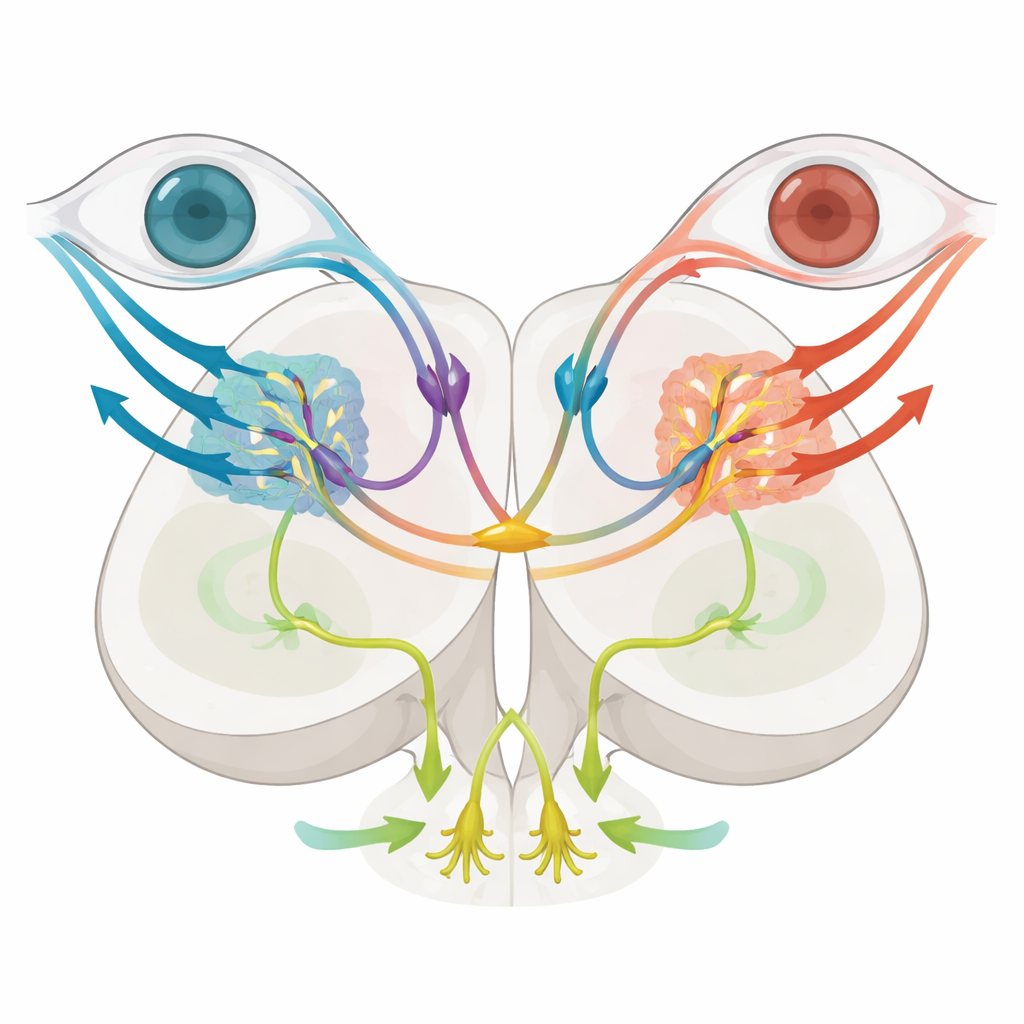
Разделение ответов глаз и тела
Для проверки функции команда селективно уничтожила nkx1.2lb-положительные претектальные нейроны, используя генетический «суицидный» фермент, который становится токсичным только в этих клетках при введении рыбкам безвредного препарата. После абляции личинки по-прежнему демонстрировали нормальные слежащие движения глаз, когда окружающий мир поворачивался вокруг них, что свидетельствует о сохранности ротационного оптического потока и вызванного им оптокинетического ответа. Однако при перемещении узора по поверхности, имитирующем движение вперед, оптомоторный ответ оказался существенно ослаблен: рыбы проходили меньшие расстояния, и их удары хвостом стали менее скоординированными, что указывает на нарушение вычисления направления плавания. Мозговая визуализация показала сокращение числа клеток, специфичных для трансляционного оптического потока, тогда как базовые монокулярные детекторы движения остались в основном сохранными. В совокупности эти эксперименты предполагают, что комиссуральные nkx1.2lb-нейроны необходимы для интеграции сигналов от обоих глаз для оценки движения прямо вперед и управления продвижением, но не требуются для стабилизации взгляда при повороте.
Почему это важно для понимания восприятия движения
Для неспециалиста ключевая мысль такова: мозг разделяет «я ли поворачиваюсь?» и «я ли двигаюсь вперед?» путем распределения этих вопросов между разными наборами клеток, даже в крошечной рыбе. Авторы показывают, что специфический ингибирующий мост между левой и правой сторонами претектума критически важен для распознавания трансляционного движения и управления телом, тогда как другие цепи могут справляться с ротационным движением и движениями глаз без этого моста. Метод CaMPARI-seq — использование света для пометки активных клеток с последующим чтением их молекулярной идентичности — предоставляет мощный способ связать то, что делают нейроны, как они устроены и какие поведения контролируют. Выводы из этой компактной системы зебрафиш могут помочь прояснить, как более крупные мозги позвоночных, включая наш собственный, поддерживают стабильность зрения и точность навигации в движущемся мире.
Цитирование: Matsuda, K., Wang, CH., Kakinuma, H. et al. Molecular and functional dissection using CaMPARI-seq reveals the neuronal organization for dissociating optic flow-dependent behaviors. Nat Commun 17, 3411 (2026). https://doi.org/10.1038/s41467-026-71371-6
Ключевые слова: оптический поток, зебрафиш, претектум, секвенирование РНК одиночных клеток, обработка визуального движения