Clear Sky Science · pt
Nova modificação proteolítica pós‑translacional no canal de potássio dependente de voltagem KCNQ2
Por que pequenas portas cerebrais importam
Nossos pensamentos, movimentos e até os primeiros choros dos recém‑nascidos dependem de bilhões de neurônios disparando em padrões temporais precisos. Esses sinais elétricos são controlados por “portas” microscópicas na membrana celular que permitem a entrada e saída de partículas carregadas. Quando essas portas falham, o resultado pode ser epilepsia em bebês recém‑nascidos. Este estudo investiga uma dessas portas, um canal de potássio chamado KCNQ2, e revela uma maneira até então desconhecida pela qual a célula parece recortar essa proteína em pedaços — um processo que pode ajudar a explicar por que algumas alterações no gene KCNQ2 causam crises breves e benignas enquanto outras levam a problemas cerebrais graves e persistentes.
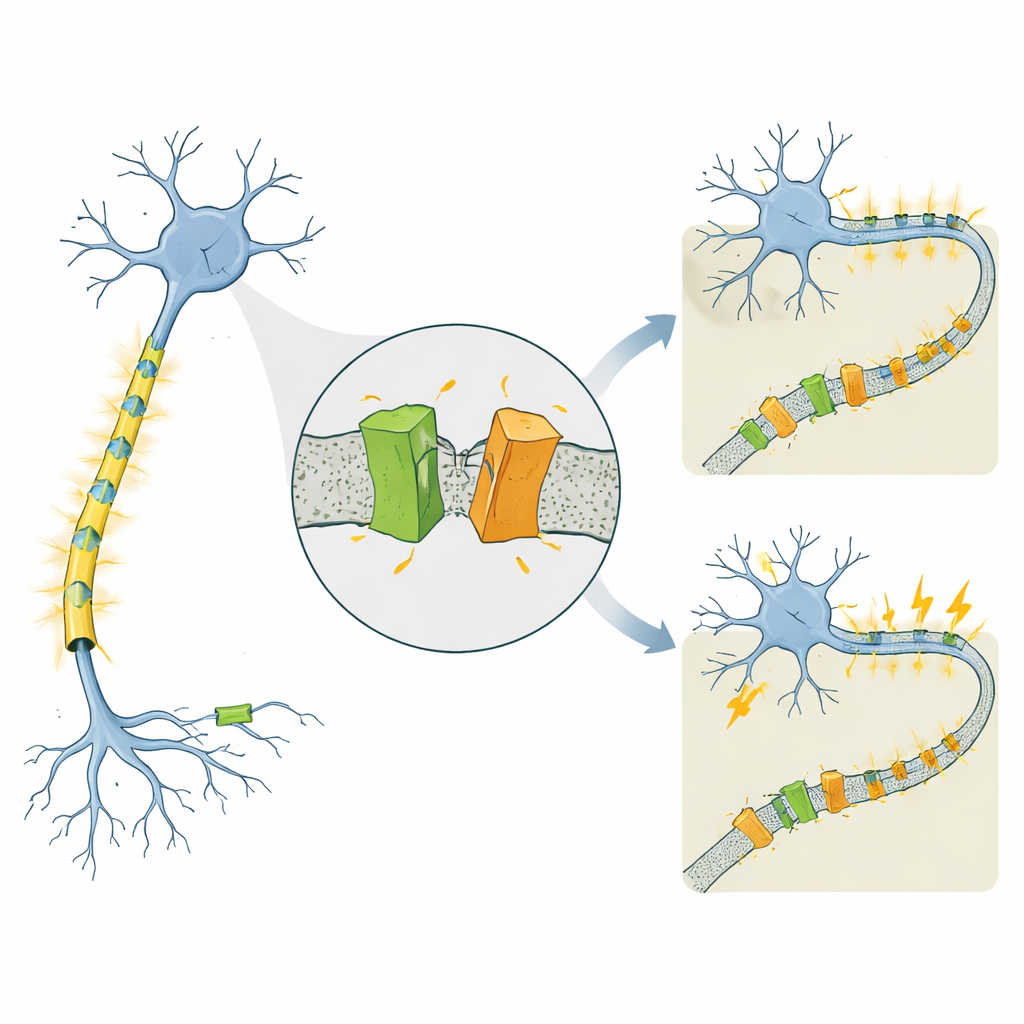
Um canal ligado a epilepsias muito distintas
KCNQ2 pertence a uma família de canais de potássio dependentes de voltagem que atuam como freios nos neurônios, ajudando-os a se recompor após cada pulso elétrico. Mutações no gene humano KCNQ2 são conhecidas por causar duas condições marcadamente diferentes que se iniciam nos primeiros dias de vida. Na epilepsia neonatal familiar autolimitada (SLFNE), as crises geralmente cessam em algumas semanas e as crianças têm bom prognóstico. Na encefalopatia epiléptica e do desenvolvimento (DEE), as crises são mais difíceis de controlar e vêm acompanhadas de atrasos graves no desenvolvimento. Muitas mutações patogênicas do KCNQ2 já haviam sido catalogadas, mas permanecia obscuro por que algumas mudanças resultam em um curso relativamente benigno enquanto outras produzem incapacidade severa, especialmente porque os níveis totais da proteína KCNQ2 frequentemente parecem semelhantes.
Um corte surpreendente no canal
Os pesquisadores estudaram versões murinas e humanas do KCNQ2, focando em várias mutações derivadas de pacientes. Eles marcaram a proteína para poder visualizar tanto sua extremidade N quanto sua extremidade C em géis laboratoriais. Como esperado, observaram o canal de comprimento total, que se localiza na membrana celular. Mas também detectaram de forma consistente duas porções menores: uma proveniente da ponta dianteira da proteína e outra da cauda. Ambos os fragmentos foram encontrados na fração rica em membrana da célula, indicando que o canal completo é formado primeiro e depois cortado, em vez de ser traduzido desde o início como formas curtas. Isso aponta para um evento de “poda” pós‑translacional até então não reconhecido — essencialmente, uma tesoura protéica da célula que divide o KCNQ2 em dois fragmentos ancorados à membrana.
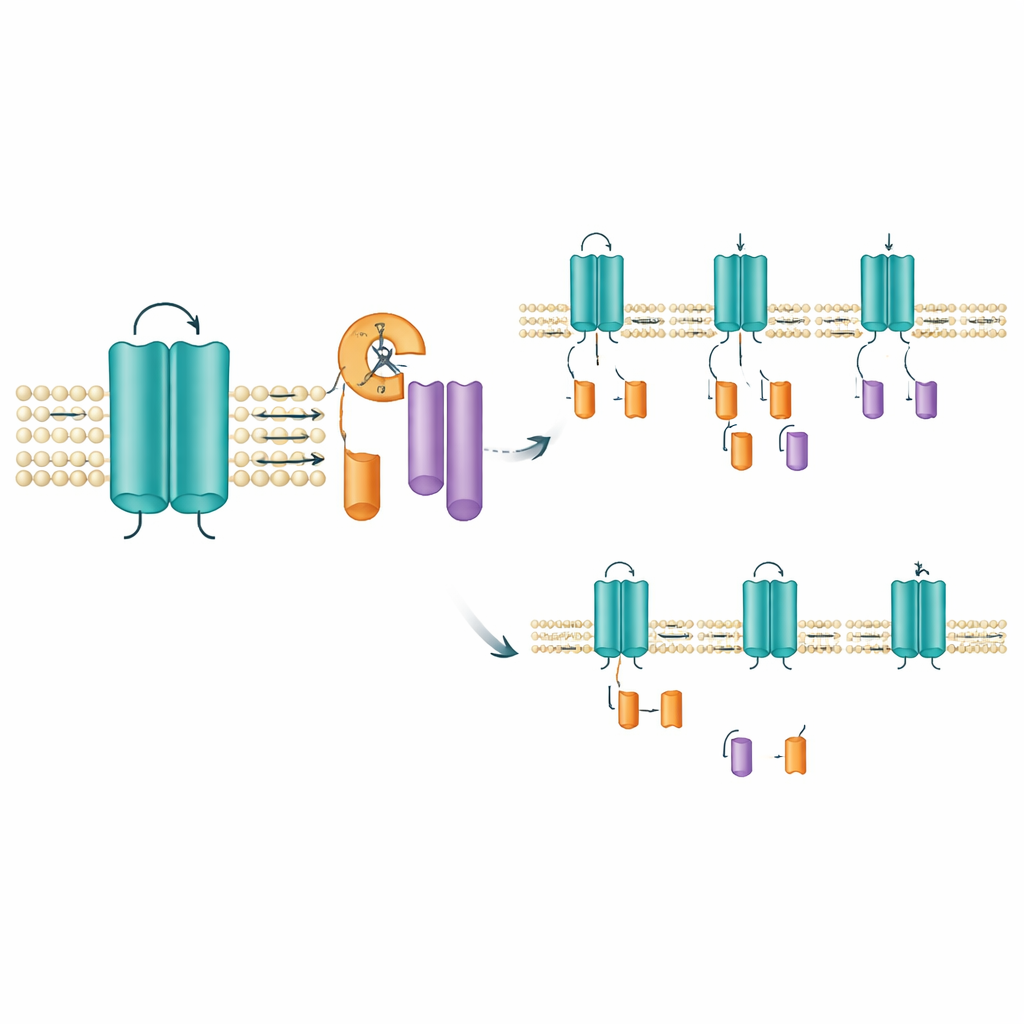
Localizando onde e como o corte ocorre
Para identificar o provável sítio de clivagem, a equipe deletou pequenos trechos de aminoácidos dentro da região que atravessa um dos segmentos sensoriais de voltagem do canal. Remover uma janela estreita de dez aminoácidos (posições 171–180) aboliu o aparecimento do fragmento da cauda enquanto deixava o canal de comprimento total intacto, sugerindo fortemente que a clivagem ocorre nessa faixa curta ou nas proximidades. Essa área situa‑se dentro de um segmento transmembrana que ajuda o canal a responder a mudanças de voltagem, de modo que um corte aí pode alterar como o canal se comporta ou quanto tempo permanece na membrana. Intrigantemente, ferramentas padrão para prever onde enzimas digestivas conhecidas cortam proteínas, assim como inibidores de proteases de amplo espectro, não impediram a formação dos fragmentos. Isso levanta a possibilidade de que uma enzima especializada embutida na membrana, do tipo já conhecido por aparar outras proteínas de sinalização, possa ser a responsável.
Mutações deslocam o equilíbrio de fragmentos
Nem todas as mutações em KCNQ2 afetaram os fragmentos da mesma maneira. Em células que expressavam mutações associadas à forma mais branda SLFNE, como A306T e uma alteração específica na posição 284 (Y284C), a quantidade do fragmento da cauda aumentou em comparação com o canal normal. Por outro lado, várias mutações ligadas à DEE, incluindo uma alteração diferente na mesma posição (Y284D), produziram menos fragmentos. Esses deslocamentos opostos — mais fragmentos em algumas variantes, menos em outras — ocorreram mesmo que a quantidade total de canal de comprimento total fosse semelhante. O mesmo padrão apareceu quando a equipe examinou a versão humana do KCNQ2 e ao testar diferentes tipos celulares, incluindo células de aparência nervosa humanas. Um canal relacionado, KCNQ3, não mostrou essa clivagem, enfatizando que esse aparo é uma característica específica e conservada do KCNQ2.
O que isso significa para a epilepsia
Este trabalho revela que o KCNQ2 não é simplesmente produzido e inserido na membrana como uma única peça estática. Em vez disso, as células rotineiramente o cortam em dois fragmentos ancorados à membrana por um processo ainda misterioso, e mutações causadoras de doença alteram quanto do canal fica na forma completa versus na forma cortada. Embora o papel exato desses fragmentos ainda não seja conhecido, sua conservação entre espécies e tipos celulares sugere que fazem parte do controle normal do canal, e não meros artefatos de laboratório. Diferenças sutis em quão eficientemente o KCNQ2 é clivado — e em como os fragmentos resultantes influenciam a atividade do canal ou sua estabilidade — podem ajudar a explicar por que algumas alterações genéticas levam a crises breves e autolimitadas enquanto outras predispõem a epilepsia grave e persistente. Estudos futuros que identifiquem a enzima responsável pelo corte e testem como os fragmentos afetam a função do canal podem abrir novos caminhos para tratamentos direcionados da epilepsia.
Citação: Kimura, Y., Uchiyama, H., Masuda, K. et al. Novel proteolytic post-translational modification in voltage-gated potassium channel KCNQ2. Sci Rep 16, 11954 (2026). https://doi.org/10.1038/s41598-026-42444-9
Palavras-chave: KCNQ2, canal de potássio, epilepsia, modificação pós‑translacional, clivagem proteolítica