Clear Sky Science · pl
AcrIIA7 przechwytuje tracrRNA, by zablokować system CRISPR‑Cas
Jak wirusy przechytrzają potężne narzędzie do edycji genów
CRISPR–Cas9 jest znany jako narzędzie do edycji genów, ale w przyrodzie pełni rolę systemu ochronnego, którego bakterie używają do cięcia obcej wirusowej DNA. To badanie ujawnia, w jaki sposób wirusowy kontratak — małe białko zwane AcrIIA7 — wyłącza tę bakteryjną obronę w nieoczekiwany sposób. Zrozumienie tego wirusowego chwyru pogłębia naszą wiedzę o mikroskopijnej rywalizacji bakterii i wirusów i wskazuje nowe sposoby włączania i wyłączania narzędzi CRISPR w laboratorium i klinice.
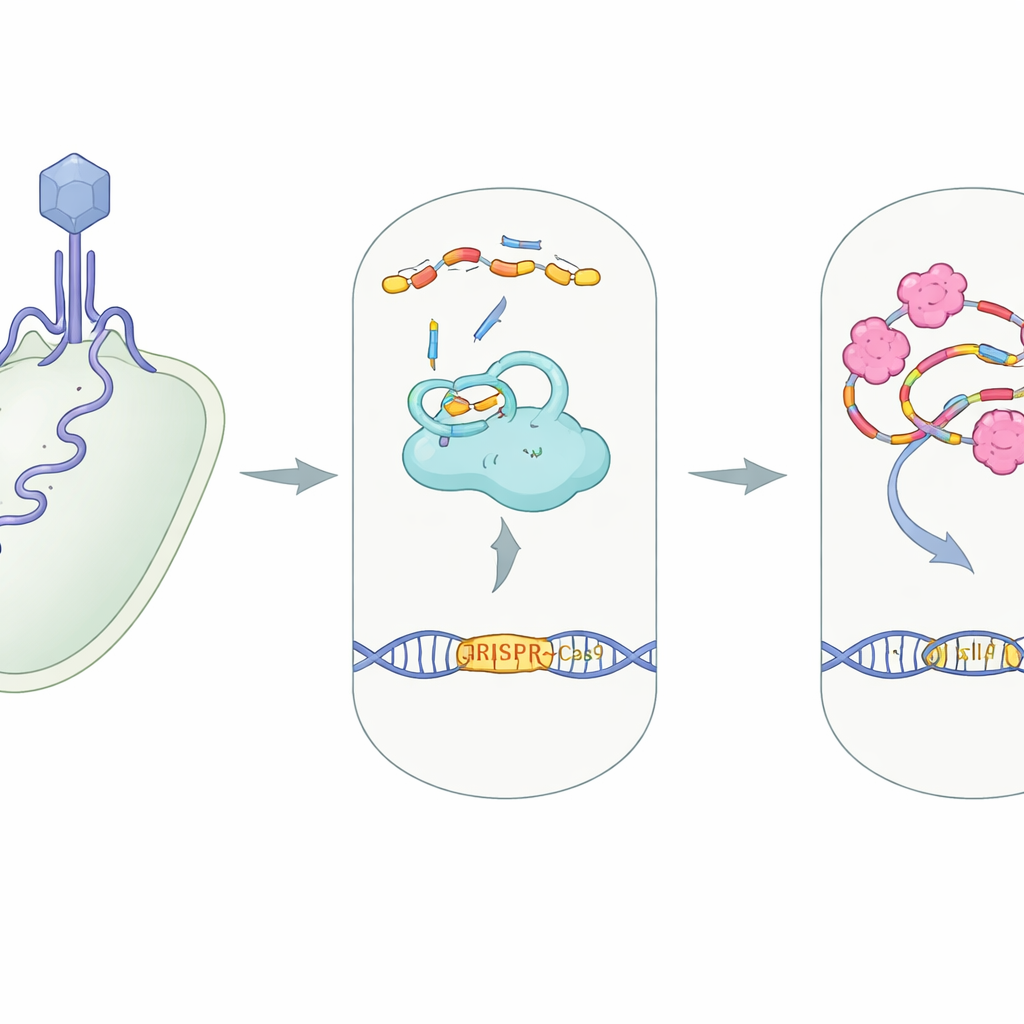
Typowa rola CRISPR jako komórkowego systemu ochronnego
W wielu bakteriach CRISPR–Cas9 działa jak molekularna kamera nadzorująca połączona z nożycami. Duże białko Cas9 współpracuje z dwoma małymi odcinkami RNA — crRNA i tracrRNA — które łączą się w kompleks prowadzący. Ten kompleks kieruje Cas9 do dowolnej sekwencji DNA pasującej do przewodnika, często materiału genetycznego obcego wirusa, aby Cas9 mógł go przeciąć i unieszkodliwić. Zbudowanie tej maszyny białkowo‑RNA, znanej jako kompleks RNP, jest kluczowym krokiem: bez prawidłowego dopasowania RNA przewodników do Cas9 system nie może rozpoznać ani przeciąć DNA intruza.
Wirusowi sabotażyści wymierzeni w CRISPR
Wirusy atakujące bakterie, zwane bakteriofagami, wyewoluowały „anty‑CRISPR” — białka sabotażujące tę obronę na niemal każdym etapie, od blokowania wiązania DNA po zatykanie miejsc cięcia Cas9. AcrIIA7 należy do rodziny znanej z ukierunkowania na Cas9, ale dokładny mechanizm jego działania był niejasny. Autorzy skupili się na AcrIIA7 z bakterii jelitowej Phocaeicola dorei, określając jego trójwymiarową strukturę i badając zachowanie w roztworze. Odkryli, że cztery cząsteczki AcrIIA7 łączą się w zwartą tetramerową strukturę złożoną z trzech domen, w tym elastycznego „głównego” regionu. Powierzchnia tego tetrameru ma głęboką, dodatnio naładowaną szczelinę — zachęcającą kieszeń dla ujemnie naładowanych kwasów nukleinowych, takich jak RNA.
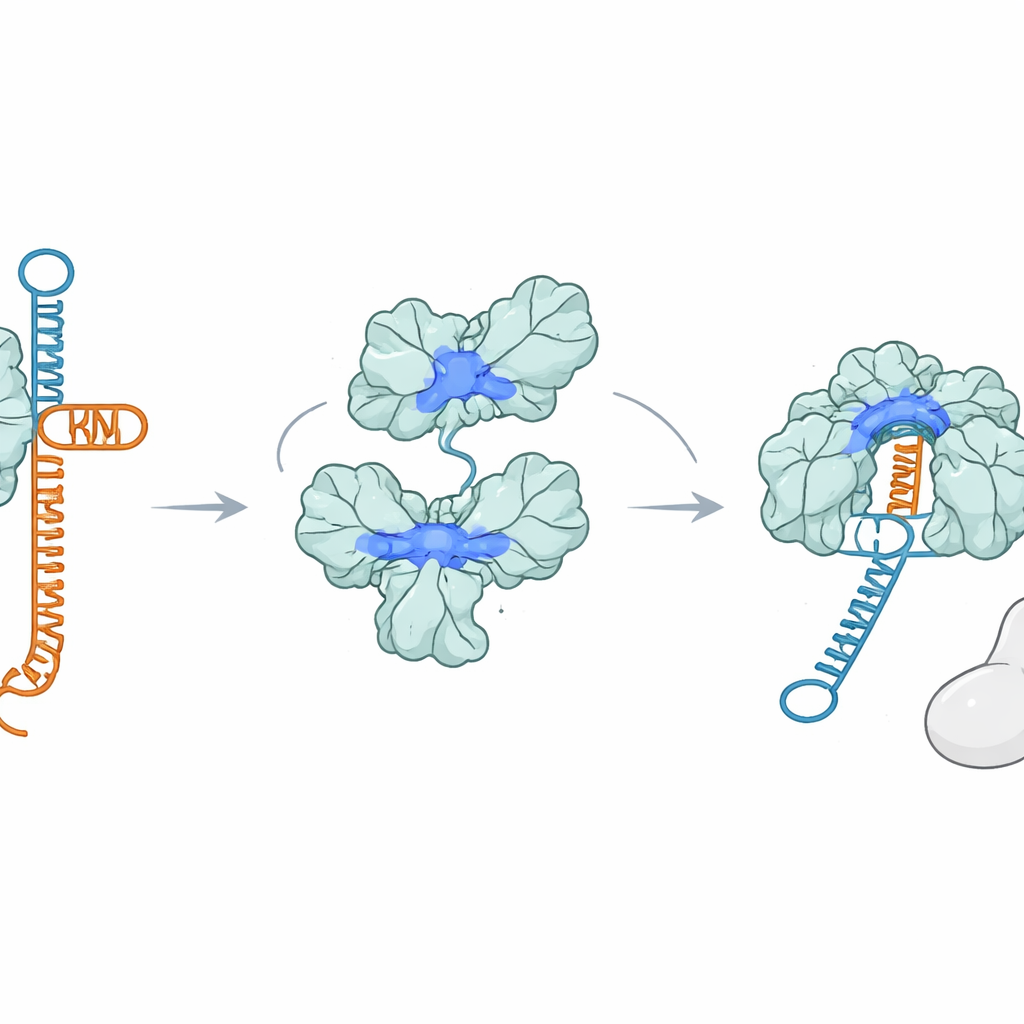
Białko, które chwyta RNA zamiast Cas9
Co zaskakujące, AcrIIA7 wcale nie wiąże się z Cas9. Wykorzystując kilka testów wiązania, badacze nie zaobserwowali stabilnej interakcji między AcrIIA7 a Cas9 ani z żadną z pojedynczych części Cas9. Zamiast tego stwierdzili, że AcrIIA7 wiąże się bezpośrednio i selektywnie z RNA. Ignorowało ono DNA jedno‑ i dwuniciowe, ale mocno przyłączało się do tracrRNA, jednej z dwóch cząsteczek RNA, które zwykle parują się, tworząc przewodnik. AcrIIA7 rozpoznawało nie tyle dokładne litery RNA, co specyficzny sfałdowany kształt złożony z dwóch sąsiadujących pętli typu hairpin. Jeśli te pętle zostały usunięte lub zmieniono ogólną architekturę łodygi‑pętli, wiązanie osłabiało się lub znikało. To wykazało, że AcrIIA7 czyta strukturę RNA bardziej niż jego sekwencję.
Przechwycenie przewodnika zanim powstanie
Przez zaciskanie się wokół tracrRNA, AcrIIA7 uniemożliwia tracrRNA parowanie się z crRNA i złożenie przewodnika potrzebnego Cas9. Eksperymenty in vitro pokazały, że gdy AcrIIA7 zetknie się z tracrRNA przed Cas9, dalsze kroki zatrzymują się: pełny RNA przewodnik formuje się słabo, a Cas9 nie może skutecznie ciąć docelowego DNA. Jednak gdy kompletny kompleks Cas9–RNA jest już zmontowany, AcrIIA7 nie jest w stanie go wypchnąć, co podkreśla, że jego główne działanie ma miejsce wcześnie, podczas formowania przewodnika. Mutacje dodatnio naładowanych aminokwasów wyściełających szczelinę tetrameru osłabiły zarówno wiązanie RNA, jak i hamowanie Cas9, co wspiera hipotezę, że tracrRNA zagnieżdża się w tej szczelinie podczas przechwycenia przez AcrIIA7.
Dwie wersje tego samego sabotażysty
Zespół porównał także pełnej długości AcrIIA7, zawierające elastyczną domenę głowy, z krótszą, naturalnie występującą wersją pozbawioną tego regionu. Obie formy tworzą tetramery i potrafią wiązać tracrRNA, a nawet małe cykliczne cząsteczki sygnałowe używane w innych ścieżkach odpornościowych. Tylko jednak wariant bez domeny głowy silnie blokuje Cas9, gdy obecny jest w pełni uformowany RNA przewodnik, co sugeruje, że region głowy częściowo przesłania dostęp do dłuższych struktur RNA. W rezultacie białko pełnej długości głównie przeszkadza w najwcześniejszym etapie — sekwestrując wolne tracrRNA — podczas gdy krótsza forma może także skuteczniej wiązać i blokować dojrzały kompleks przewodnika.
Co to oznacza dla biologii i edycji genów
Dla niespecjalisty kluczowy przekaz jest taki, że to wirusowe białko dezaktywuje CRISPR nie przez bezpośredni atak na słynne „nożyce” Cas9, lecz przez kradzież jego szkieletu RNA zanim maszyna tnąca zostanie zbudowana. To ujawnia nowy słaby punkt w obronie CRISPR: kontakty RNA–RNA i RNA–białko konieczne do złożenia działającego kompleksu. W naturze taka strategia pomaga wirusom wymknąć się przed bakteryjną odpornością. W laboratorium molekuły inspirowane AcrIIA7 mogłyby oferować precyzyjne, odwracalne sposoby wyłączania narzędzi CRISPR przez celowanie w ich przewodnikowe RNA, poprawiając bezpieczeństwo i kontrolę przyszłych terapii edycji genomu.
Cytowanie: Lee, S.Y., Park, H.H. AcrIIA7 hijacks tracrRNA to block CRISPR-Cas system. Nat Commun 17, 3959 (2026). https://doi.org/10.1038/s41467-026-70749-w
Słowa kluczowe: CRISPR‑Cas9, białka anty‑CRISPR, tracrRNA, bakteriofag, kontrola edycji genów