Clear Sky Science · nl
AcrIIA7 kaapt tracrRNA om CRISPR-Cas-systeem te blokkeren
Hoe virussen een krachtig genbewerkingsgereedschap te slim af zijn
CRISPR–Cas9 is beroemd als gereedschap voor genbewerking, maar in de natuur functioneert het als een beveiligingssysteem dat bacteriën gebruiken om binnendringend viraal DNA in stukjes te hakken. Deze studie onthult hoe een viraal tegenwapen, een klein eiwit genaamd AcrIIA7, die bacteriële verdediging op een onverwachte manier uitschakelt. Begrijpen hoe dit virale trucje werkt verdiept niet alleen ons inzicht in de microscopische wapenwedloop tussen bacteriën en virussen, het wijst ook op nieuwe manieren om CRISPR-gereedschap in het laboratorium en de kliniek aan- en uit te schakelen.
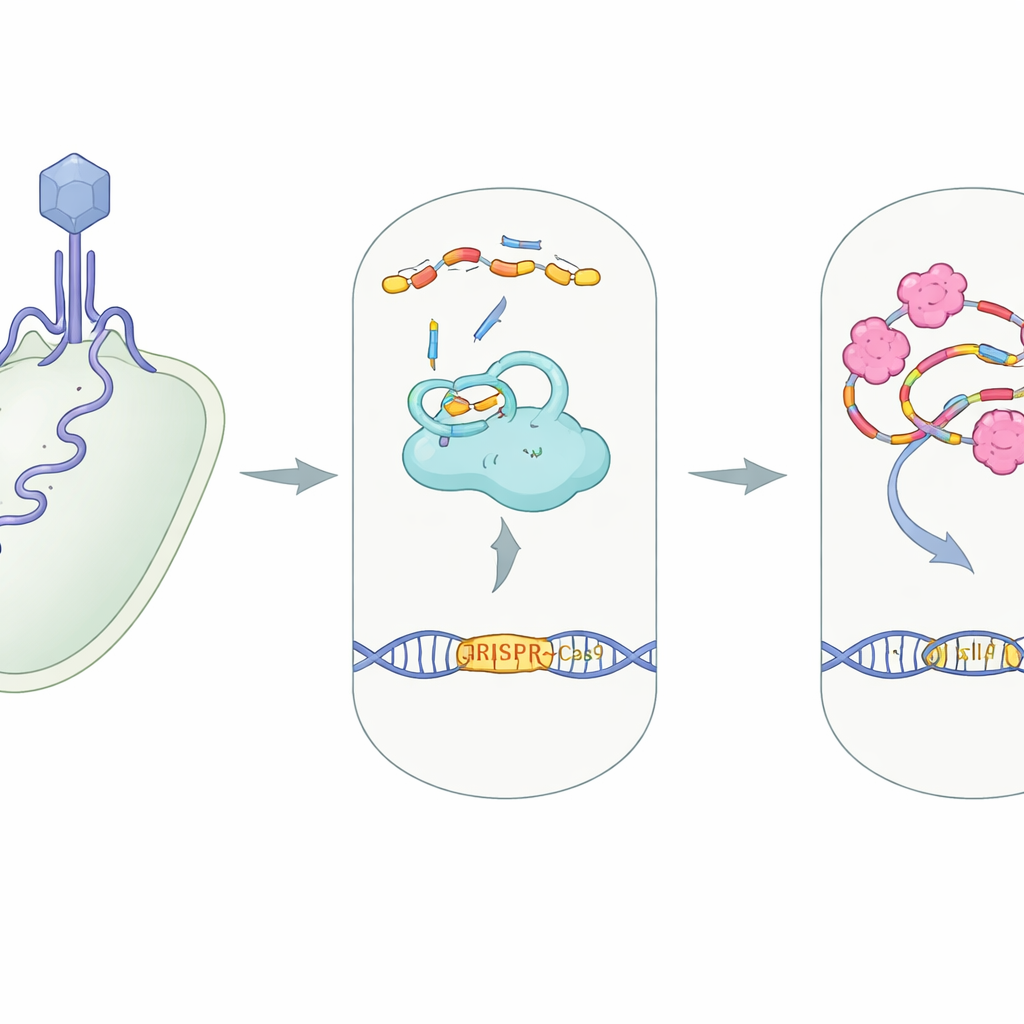
De gebruikelijke rol van CRISPR als cellulaire beveiliging
In veel bacteriën werkt CRISPR–Cas9 als een moleculaire bewakingscamera gekoppeld aan een schaar. Een groot eiwit, Cas9, werkt samen met twee kleine RNA-strengen—crRNA en tracrRNA—die in elkaar klikken tot een gidscomplex. Dit complex leidt Cas9 naar elk DNA-segment dat overeenkomt met de gids, vaak het genetisch materiaal van een binnendringend virus, zodat Cas9 het kan knippen en neutraliseren. Het bouwen van dit eiwit–RNA-machientje, bekend als een RNP-complex, is een essentiële stap: zonder dat de gids-RNA’s correct in Cas9 passen, kan het systeem de indringer niet herkennen of knippen.
Virale saboteurs gericht op CRISPR
Virussen die bacteriën infecteren, bacteriofagen genoemd, hebben "anti‑CRISPR"-eiwitten ontwikkeld die deze verdediging op bijna elk niveau saboteert, van het blokkeren van DNA‑binding tot het blokkeren van Cas9’s knipplaatsen. AcrIIA7 behoort tot een familie die bekendstaat om Cas9 te targeten, maar precies hoe het werkte was onduidelijk. De auteurs richtten zich op AcrIIA7 uit een darmbacterie, Phocaeicola dorei, bepaalden zijn driedimensionale structuur en testten zijn gedrag in oplossing. Ze ontdekten dat vier AcrIIA7-moleculen samen een compact tetramer vormen dat uit drie domeinen bestaat, waaronder een flexibele "kop"-regio. Het oppervlak van dit tetramer heeft een diepe, positief geladen groeve—een uitnodigende holte voor negatief geladen nucleïnezuren zoals RNA.
Een eiwit dat RNA grijpt in plaats van Cas9
Verrassend genoeg hecht AcrIIA7 zich helemaal niet aan Cas9. Met verschillende bindingsproeven zagen de onderzoekers geen stabiele interactie tussen AcrIIA7 en Cas9, noch met afzonderlijke delen van Cas9. In plaats daarvan vonden ze dat AcrIIA7 direct en selectief aan RNA bindt. Het negeerde enkel- en dubbelstrengs DNA, maar hechtte sterk aan tracrRNA, een van de twee RNA’s die normaal samenvouwen om de gids te vormen. AcrIIA7 herkende niet de exacte RNA-lettervolgorde maar een specifieke gevouwen vorm bestaande uit twee aangrenzende haarspeldlussen. Als die lussen werden verwijderd of de algemene stengel‑lus‑architectuur werd gewijzigd, verzwakte of verdween de binding. Dit toonde aan dat AcrIIA7 de structuur van het RNA leest meer dan de sequentie.
De gids kapen voordat deze gevormd is
Door zich vast te klemmen aan tracrRNA voorkomt AcrIIA7 dat tracrRNA met crRNA paar vormt om de gids samen te stellen die Cas9 nodig heeft. Experimenten in buisjes toonden aan dat wanneer AcrIIA7 tracrRNA ontmoet voordat Cas9 dat doet, de daaropvolgende stappen vastlopen: het volledige gids‑RNA vormt slecht en Cas9 kan het doel-DNA niet efficiënt knippen. Zodra echter een compleet Cas9–RNA-complex is samengesteld, kan AcrIIA7 het niet meer losmaken, wat benadrukt dat zijn belangrijkste werking vroeg plaatsvindt, tijdens de gidsvorming. Het muteren van positief geladen aminozuren die de groeve van het tetramer bekleden, beschadigde zowel RNA‑binding als Cas9‑remming, wat de gedachte ondersteunt dat tracrRNA in deze groeve nestelt wanneer het door AcrIIA7 wordt gekaapt.
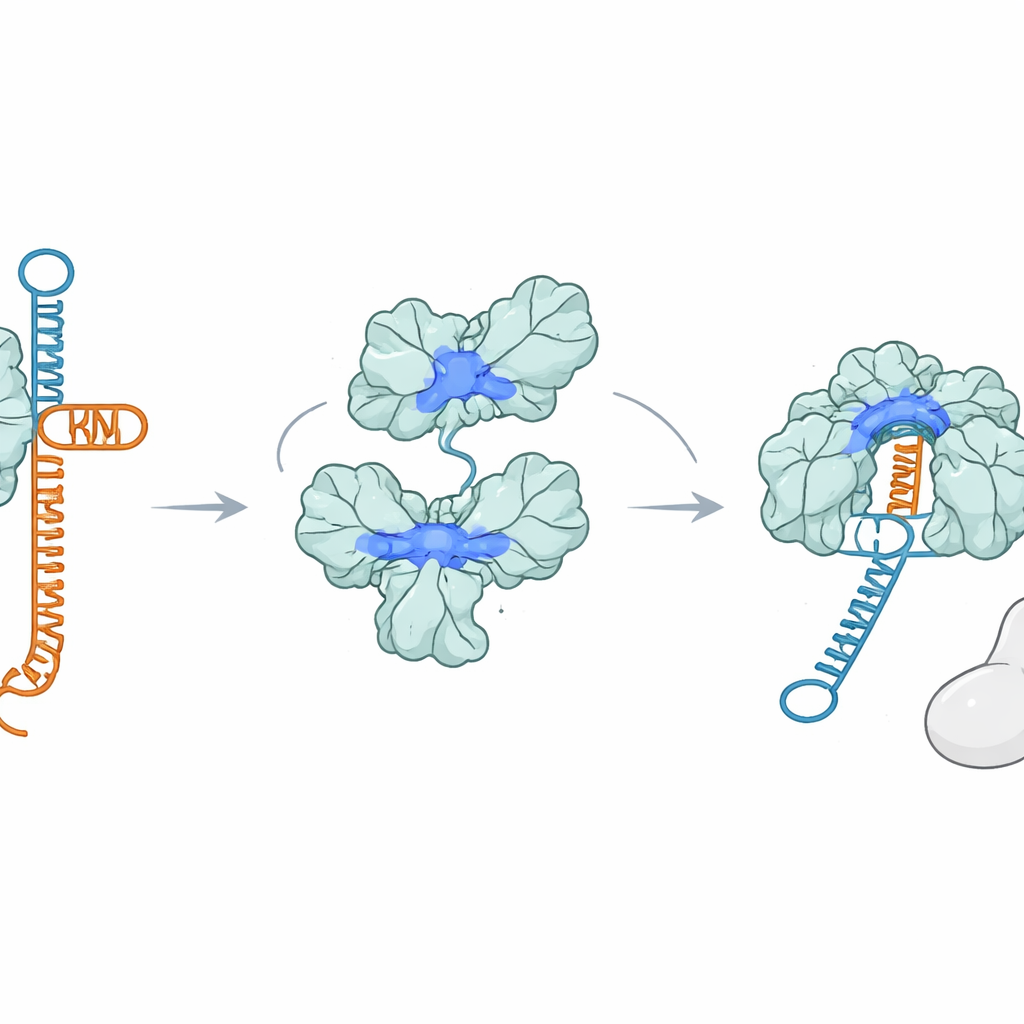
Twee versies van dezelfde saboteur
Het team vergeleek ook volledig AcrIIA7, dat het flexibele kopdomein bevat, met een kortere, van nature voorkomende variant die dit gebied mist. Beide vormen assembleren tot tetrameren en kunnen tracrRNA binden en zelfs kleine cyclische signaalmoleculen binden die in andere immuunroutes worden gebruikt. Maar slechts de koploze variant blokkeert Cas9 sterk als er een volledig gevormd gids‑RNA aanwezig is, wat suggereert dat het kopgebied de toegang tot langere RNA‑structuren deels belemmert. Als gevolg daarvan bemoeilijkt het volledige eiwit vooral de vroegste stap—het afschermen van vrij tracrRNA—terwijl de kortere vorm ook het volwassen gidscomplex effectiever kan binden en blokkeren.
Wat dit betekent voor biologie en genbewerking
Voor een niet‑specialist is de kernboodschap dat dit virale eiwit CRISPR uitschakelt niet door de beroemde Cas9‑"schaar" direct aan te vallen, maar door het RNA‑steunframe ervan te stelen voordat de knipmachine is gebouwd. Dit onthult een nieuw zwak punt in CRISPR‑verdedigingen: de RNA–RNA en RNA–eiwitinteracties die nodig zijn om een werkend complex samen te stellen. In de natuur helpt zo’n strategie virussen om langs bacteriële immuniteit te glippen. In het laboratorium zouden aan AcrIIA7 ontleende moleculen precieze, omkeerbare manieren kunnen bieden om CRISPR-gereedschap uit te schakelen door hun RNA‑gidsen te targeten, wat de veiligheid en controle van toekomstige genbewerkingstherapieën kan verbeteren.
Bronvermelding: Lee, S.Y., Park, H.H. AcrIIA7 hijacks tracrRNA to block CRISPR-Cas system. Nat Commun 17, 3959 (2026). https://doi.org/10.1038/s41467-026-70749-w
Trefwoorden: CRISPR-Cas9, anti-CRISPR-eiwitten, tracrRNA, bacteriofaag, controle van genbewerking