Clear Sky Science · it
AcrIIA7 dirotta la tracrRNA per bloccare il sistema CRISPR-Cas
Come i virus ingannano un potente strumento di editing genetico
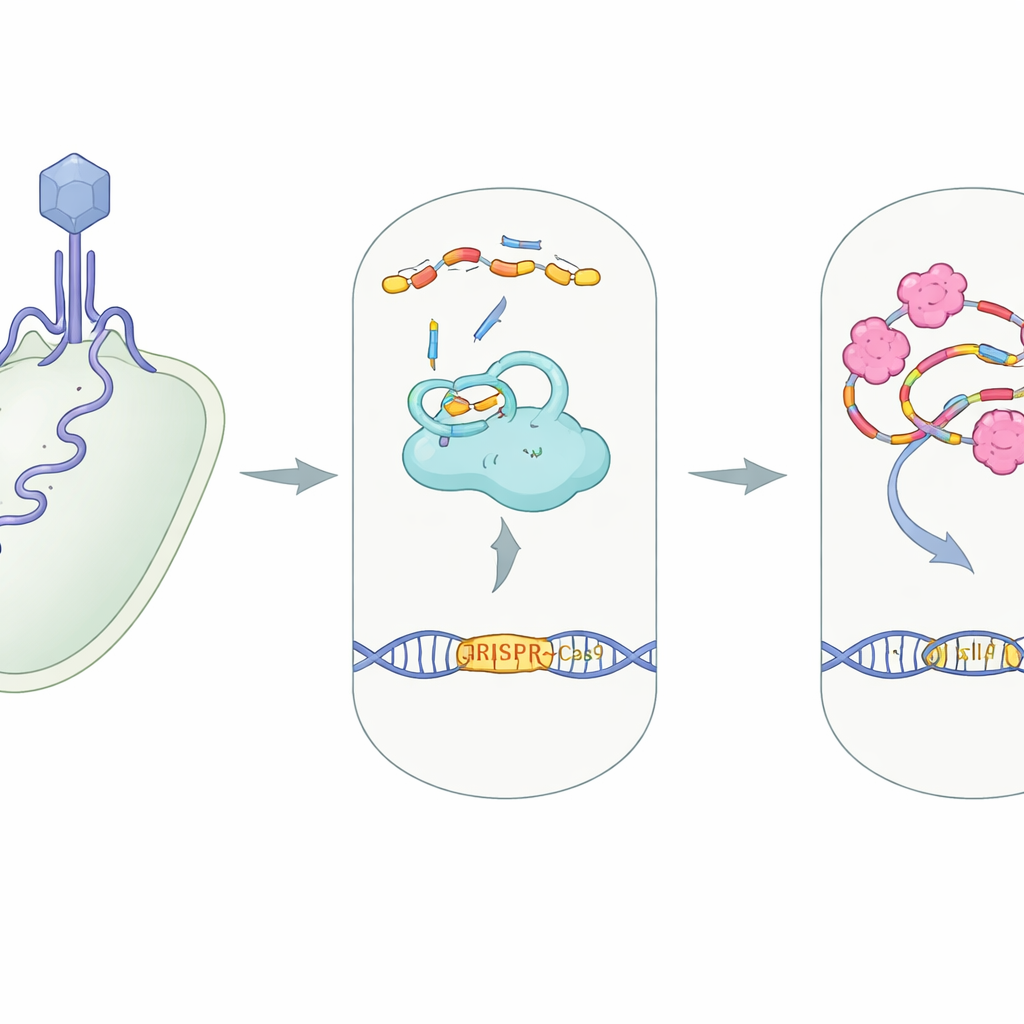
CRISPR–Cas9 è noto come strumento di editing genetico, ma in natura funziona come un sistema di sicurezza che i batteri usano per tagliare il DNA virale invasore. Questo studio rivela come un contro-arma virale, una piccola proteina chiamata AcrIIA7, disattivi quella difesa batterica in modo inaspettato. Capire questo trucco virale non solo approfondisce la nostra conoscenza della corsa agli armamenti microscopica tra batteri e virus, ma indica anche nuovi modi per accendere e spegnere gli strumenti CRISPR in laboratorio e in clinica.
Il ruolo usuale di CRISPR come sicurezza cellulare
In molti batteri, CRISPR–Cas9 funziona come una telecamera molecolare abbinata a delle forbici. Una grande proteina, Cas9, si unisce a due piccoli filamenti di RNA—crRNA e tracrRNA—che si compongono in un complesso guida. Questo complesso dirige Cas9 verso qualsiasi sequenza di DNA che corrisponda alla guida, spesso il materiale genetico di un virus invasore, in modo che Cas9 possa tagliarlo e neutralizzarlo. La costruzione di questa macchina proteina–RNA, nota come complesso RNP, è un passaggio essenziale: senza che gli RNA guida si adattino correttamente a Cas9, il sistema non può riconoscere né tagliare il DNA dell’intruso.
Sabotatori virali mirati a CRISPR
I virus che infettano i batteri, chiamati batteriofagi, hanno evoluto proteine “anti‑CRISPR” che sabotano questa difesa a quasi ogni stadio, dal bloccare il legame al DNA al inceppare i siti di taglio di Cas9. AcrIIA7 appartiene a una famiglia nota per prendere di mira Cas9, ma il suo meccanismo esatto non era chiaro. Gli autori si sono concentrati su AcrIIA7 proveniente da un batterio intestinale, Phocaeicola dorei, determinandone la struttura tridimensionale e testandone il comportamento in soluzione. Hanno scoperto che quattro molecole di AcrIIA7 si uniscono in un compatto tetramero composto da tre domini, inclusa una regione “testa” flessibile. La superficie di questo tetramero presenta una profonda scanalatura carica positivamente—una tasca invitante per acidi nucleici carichi negativamente come l’RNA.
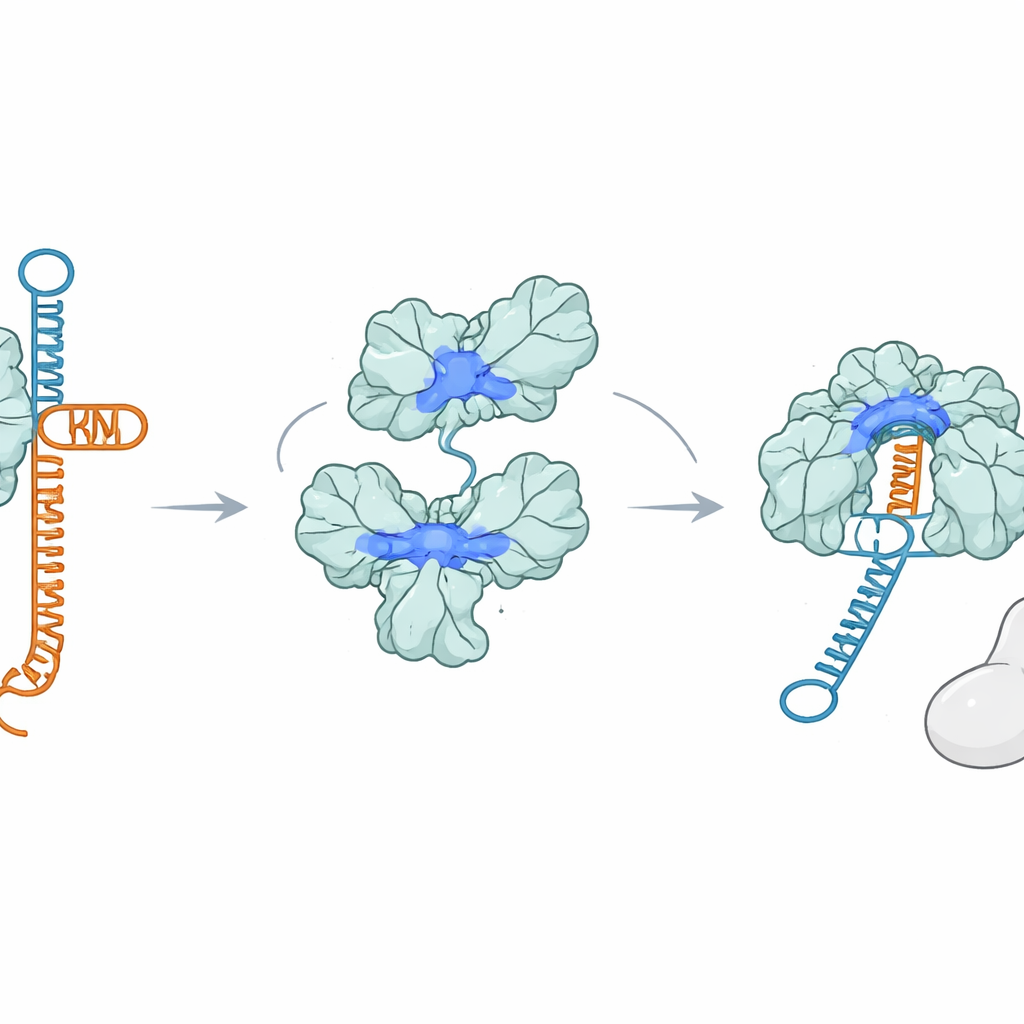
Una proteina che afferra l’RNA invece di Cas9
Sorprendentemente, AcrIIA7 non si lega affatto a Cas9. Usando diversi test di legame, i ricercatori non hanno osservato interazioni stabili tra AcrIIA7 e Cas9, né con nessuna delle singole parti di Cas9. Hanno invece scoperto che AcrIIA7 si lega direttamente e selettivamente all’RNA. Ignorava il DNA a singolo e doppio filamento, ma si legava con forza alla tracrRNA, uno dei due RNA che normalmente si appaiano per formare la guida. AcrIIA7 non riconosceva le singole lettere dell’RNA ma una specifica struttura ripiegata costituita da due anse a forcina adiacenti. Se quelle anse venivano rimosse o l’architettura complessiva dello stelo-ansa cambiata, il legame si indeboliva o spariva. Ciò dimostrava che AcrIIA7 interpreta più la struttura dell’RNA che la sequenza.
Dirottare la guida prima che si formi
Aggrappandosi alla tracrRNA, AcrIIA7 impedisce alla tracrRNA di appaiarsi con la crRNA per assemblare la guida necessaria a Cas9. Esperimenti in provetta hanno mostrato che quando AcrIIA7 incontra la tracrRNA prima di Cas9, i passaggi successivi si bloccano: la guida completa si forma male e Cas9 non può tagliare efficacemente il DNA bersaglio. Tuttavia, una volta che un complesso Cas9–RNA completo è assemblato, AcrIIA7 non riesce più a scardinarlo, sottolineando che la sua azione principale avviene precocemente, durante la formazione della guida. Mutare gli amminoacidi carichi positivamente che fiancheggiano la scanalatura del tetramero comprometteva sia il legame con l’RNA sia l’inibizione di Cas9, a supporto dell’idea che la tracrRNA si incastri in questa scanalatura quando viene dirottata da AcrIIA7.
Due versioni dello stesso sabotatore
Il team ha anche confrontato AcrIIA7 a lunghezza intera, che include il dominio testa flessibile, con una versione più corta, presente in natura, che manca di questa regione. Entrambe le forme si assemblano in tetrameri e possono legare la tracrRNA e persino piccoli molecoli ciclici segnalatori usati in altre vie immunitarie. Ma solo la variante senza testa blocca fortemente Cas9 quando è presente una guida RNA completamente formata, suggerendo che la regione testa ostruisca parzialmente l’accesso a strutture RNA più lunghe. Di conseguenza, la proteina a lunghezza intera interferisce principalmente con il primo stadio—sequestrando la tracrRNA libera—mentre la forma più corta può anche legare e bloccare il complesso guida maturo in modo più efficace.
Cosa significa per la biologia e l’editing genico
Per un non specialista, il messaggio chiave è che questa proteina virale disabilita CRISPR non attaccando direttamente le famose “forbici” Cas9, ma rubando il suo impalcatura di RNA prima che la macchina da taglio venga assemblata. Questo rivela un nuovo punto debole nelle difese CRISPR: i contatti RNA–RNA e RNA–proteina necessari per assemblare un complesso funzionante. In natura, tale strategia aiuta i virus a eludere l’immunità batterica. In laboratorio, molecole ispirate ad AcrIIA7 potrebbero offrire modi precisi e reversibili per spegnere gli strumenti CRISPR mirandone le guide RNA, migliorando la sicurezza e il controllo delle future terapie di editing del genoma.
Citazione: Lee, S.Y., Park, H.H. AcrIIA7 hijacks tracrRNA to block CRISPR-Cas system. Nat Commun 17, 3959 (2026). https://doi.org/10.1038/s41467-026-70749-w
Parole chiave: CRISPR-Cas9, proteine anti-CRISPR, tracrRNA, bacteriofago, controllo dell’editing genico