Clear Sky Science · es
AcrIIA7 secuestra tracrRNA para bloquear el sistema CRISPR-Cas
Cómo los virus burlan una potente herramienta de edición genética
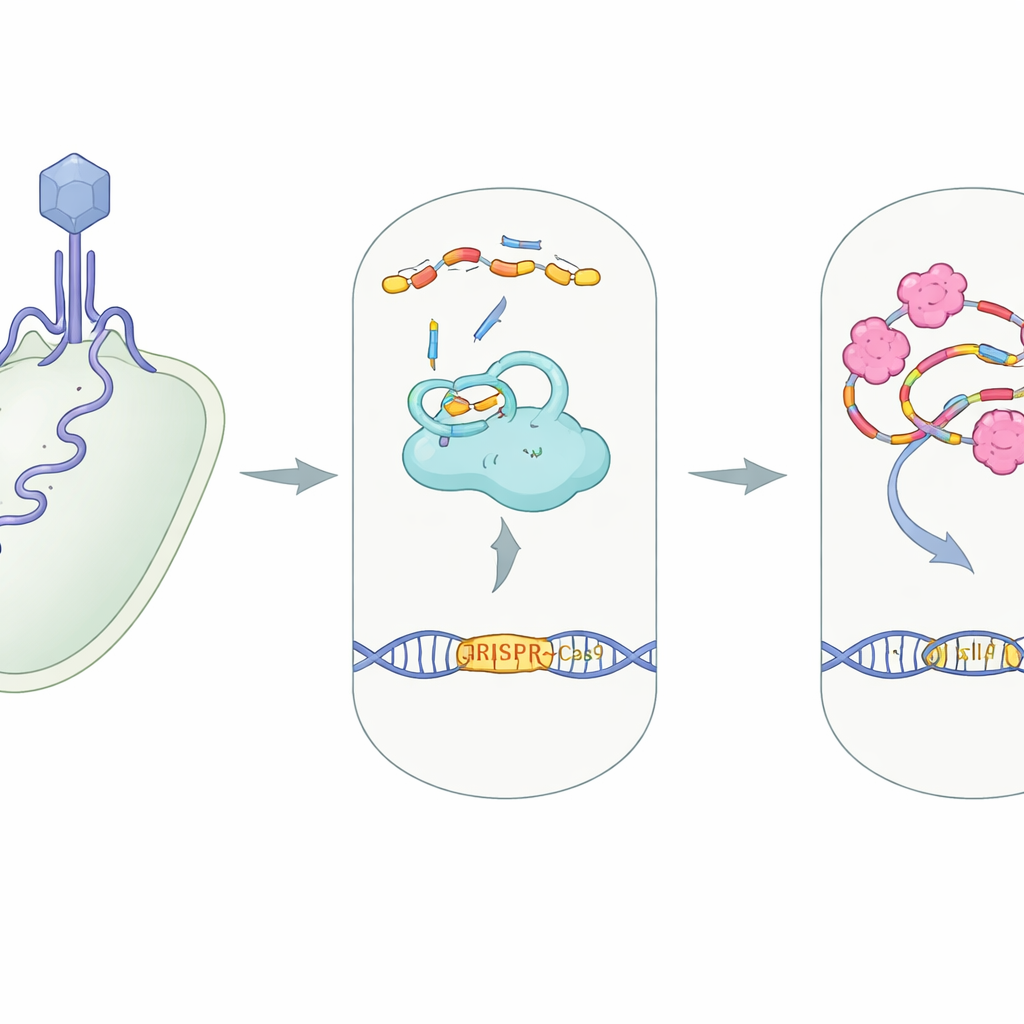
CRISPR–Cas9 es célebre como herramienta de edición genética, pero en la naturaleza funciona como un sistema de seguridad que las bacterias emplean para cortar el ADN viral invasor. Este estudio revela cómo un contramedida viral, una pequeña proteína llamada AcrIIA7, desactiva esa defensa bacteriana de una manera inesperada. Comprender este truco viral no solo profundiza nuestro conocimiento sobre la carrera armamentística microscópica entre bacterias y virus, sino que también apunta a nuevas formas de encender y apagar las herramientas CRISPR en el laboratorio y la clínica.
El papel habitual de CRISPR como seguridad celular
En muchas bacterias, CRISPR–Cas9 funciona como una cámara de vigilancia molecular equipada con tijeras. Una proteína grande, Cas9, se asocia con dos hebras pequeñas de ARN—crRNA y tracrRNA—que se ensamblan en un complejo guía. Ese complejo dirige a Cas9 hacia cualquier secuencia de ADN que coincida con la guía, a menudo el material genético de un virus invasor, para que Cas9 lo corte y neutralice. Construir esta máquina proteína‑ARN, conocida como complejo RNP, es un paso esencial: sin que los ARN guía encajen correctamente en Cas9, el sistema no puede reconocer ni cortar el ADN del intruso.
Saboteadores virales dirigidos contra CRISPR
Los virus que infectan bacterias, llamados bacteriófagos, han evolucionado proteínas “anti‑CRISPR” que sabotean esta defensa en casi todas las etapas, desde bloquear la unión al ADN hasta inutilizar los sitios de corte de Cas9. AcrIIA7 pertenece a una familia conocida por dirigirse a Cas9, pero no estaba claro cómo actuaba exactamente. Los autores se centraron en AcrIIA7 de una bacteria intestinal, Phocaeicola dorei, determinando su estructura tridimensional y probando su comportamiento en solución. Descubrieron que cuatro moléculas de AcrIIA7 se unen formando un tetramero compacto con tres dominios, incluido una región “cabeza” flexible. La superficie de este tetramero presenta una hendidura profunda y cargada positivamente—un bolsillo atractivo para ácidos nucleicos negativamente cargados como el ARN.
Una proteína que atrapa ARN en lugar de Cas9
Sorprendentemente, AcrIIA7 no se une a Cas9 en absoluto. Usando varias pruebas de unión, los investigadores no observaron interacción estable entre AcrIIA7 y Cas9, ni con ninguna de las partes individuales de Cas9. En su lugar, encontraron que AcrIIA7 se une al ARN de forma directa y selectiva. Ignoró el ADN de cadena simple y doble, pero se ligó con fuerza a la tracrRNA, uno de los dos ARN que normalmente se emparejan para formar la guía. AcrIIA7 no reconocía las letras exactas del ARN sino una conformación plegada específica formada por dos bucles en horquilla adyacentes. Si esos bucles se eliminaban o se alteraba la arquitectura general del tallo‑bucle, la unión se debilitaba o desaparecía. Esto mostró que AcrIIA7 lee más la estructura del ARN que su secuencia.
Secuestrar la guía antes de que se forme
Al aferrarse a la tracrRNA, AcrIIA7 impide que la tracrRNA se empareje con la crRNA para ensamblar la guía que necesita Cas9. Experimentos en tubo de ensayo mostraron que cuando AcrIIA7 encuentra la tracrRNA antes que Cas9, los pasos posteriores se detienen: la guía completa de ARN se forma pobremente y Cas9 no puede cortar eficientemente el ADN objetivo. Sin embargo, una vez que se ha ensamblado un complejo Cas9–ARN completo, AcrIIA7 ya no puede desalojarlo, lo que subraya que su acción principal ocurre temprano, en la formación de la guía. Mutar aminoácidos cargados positivamente que recubren la hendidura del tetramero incapacita tanto la unión al ARN como la inhibición de Cas9, lo que respalda la idea de que la tracrRNA encaja en esa hendidura cuando es secuestrada por AcrIIA7.
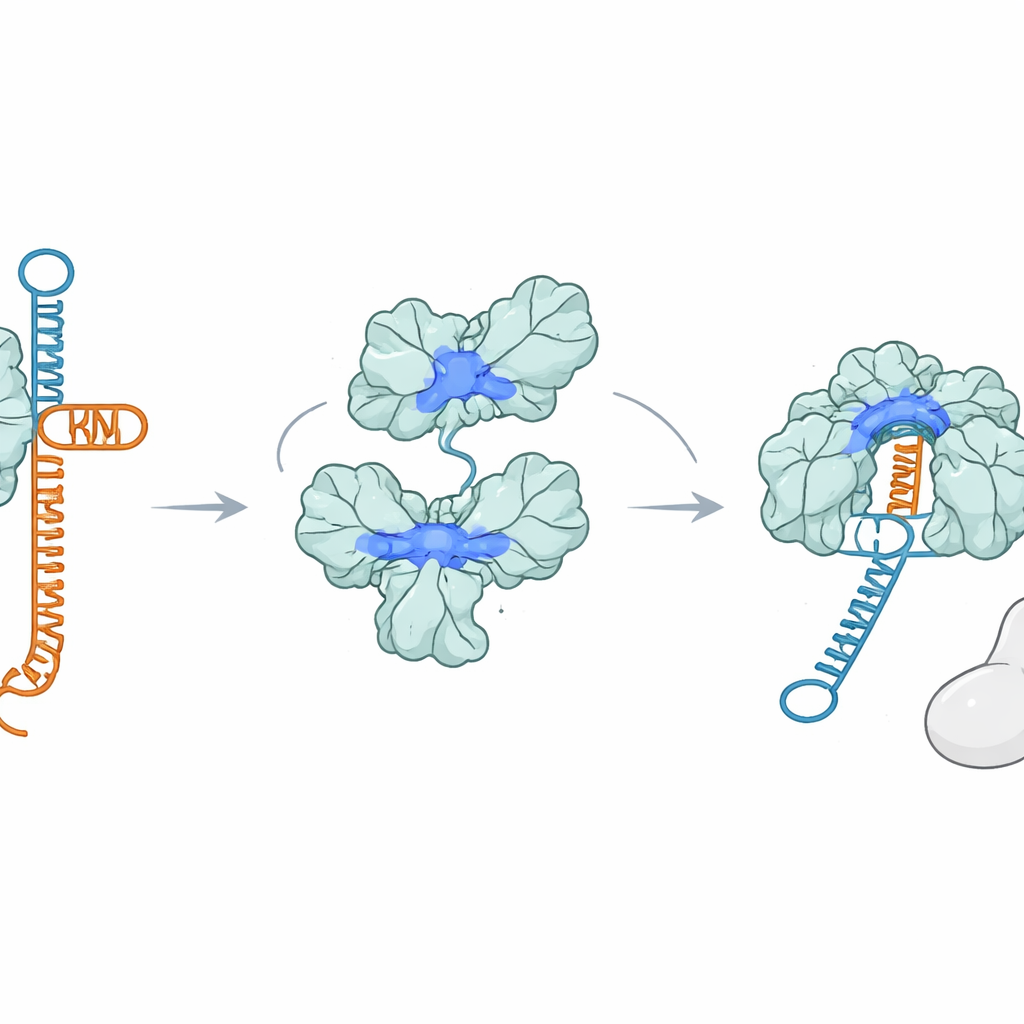
Dos versiones del mismo saboteador
El equipo también comparó AcrIIA7 de longitud completa, que incluye el dominio de cabeza flexible, con una versión más corta y natural que carece de esa región. Ambas formas se ensamblan en tetrameros y pueden unirse a la tracrRNA e incluso a pequeñas moléculas cíclicas de señalización usadas en otras vías inmunitarias. Pero solo la variante sin cabeza bloquea con fuerza a Cas9 cuando hay una guía ARN completamente formada, lo que sugiere que la región de la cabeza obstruye en parte el acceso a estructuras ARN más largas. Como resultado, la proteína de longitud completa interfiere principalmente con el paso más temprano—secuestrar la tracrRNA libre—mientras que la forma más corta también puede unirse y bloquear el complejo guía maduro con mayor eficacia.
Qué significa esto para la biología y la edición génica
Para un público no especializado, el mensaje clave es que esta proteína viral desactiva CRISPR no atacando directamente la famosa “tijera” Cas9, sino robando su andamiaje de ARN antes de que la máquina de cortar se construya. Esto revela un nuevo punto débil en las defensas CRISPR: los contactos ARN‑ARN y ARN‑proteína necesarios para ensamblar un complejo funcional. En la naturaleza, tal estrategia ayuda a los virus a eludir la inmunidad bacteriana. En el laboratorio, moléculas inspiradas en AcrIIA7 podrían ofrecer maneras precisas y reversibles de apagar herramientas CRISPR atacando sus guías de ARN, mejorando la seguridad y el control de futuras terapias de edición del genoma.
Cita: Lee, S.Y., Park, H.H. AcrIIA7 hijacks tracrRNA to block CRISPR-Cas system. Nat Commun 17, 3959 (2026). https://doi.org/10.1038/s41467-026-70749-w
Palabras clave: CRISPR-Cas9, proteínas anti-CRISPR, tracrRNA, bacteriófago, control de la edición génica