Clear Sky Science · de
AcrIIA7 kapert tracrRNA, um das CRISPR‑Cas‑System zu blockieren
Wie Viren ein mächtiges Gentechnik‑Werkzeug austricksen
CRISPR–Cas9 ist als Werkzeug zur Genbearbeitung bekannt, erfüllt in der Natur jedoch eine Schutzfunktion: Bakterien nutzen es, um eindringende virale DNA zu zerschneiden. Diese Studie zeigt, wie eine virale Gegenwaffe, ein kleines Protein namens AcrIIA7, diese bakterielle Abwehr auf überraschende Weise ausschaltet. Das Verständnis dieses viralen Tricks vertieft nicht nur unsere Einsicht in das mikroskopische Wettrüsten zwischen Bakterien und Viren, sondern weist auch auf neue Möglichkeiten hin, CRISPR‑Werkzeuge im Labor und in der Klinik ein‑ und auszuschalten.
Die übliche Rolle von CRISPR als zelluläre Sicherheitsinstanz
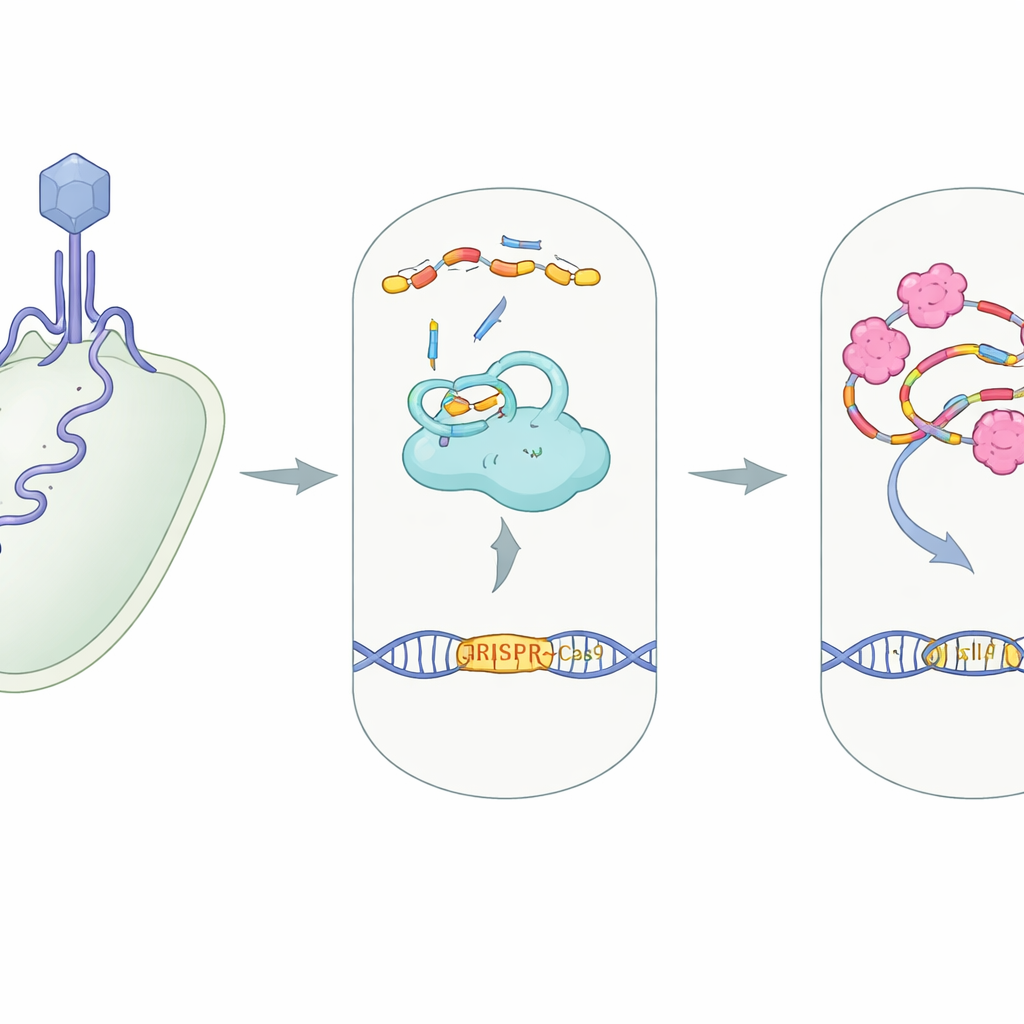
In vielen Bakterien arbeitet CRISPR–Cas9 wie eine molekulare Überwachungskamera mit Schere. Ein großes Protein, Cas9, verbindet sich mit zwei kleinen RNA‑Strängen — crRNA und tracrRNA — die zusammen ein Leitkomplex bilden. Dieser Komplex steuert Cas9 zu jeder DNA‑Sequenz, die zur Anleitung passt, häufig das Erbgut eines eindringenden Virus, sodass Cas9 es schneiden und neutralisieren kann. Der Aufbau dieser Protein‑RNA‑Maschinerie, eines sogenannten RNP‑Komplexes, ist ein wesentlicher Schritt: Ohne korrekt ineinandergreifende Leit‑RNAs kann das System die eindringende DNA weder erkennen noch zerschneiden.
Virkliche Saboteure, die auf CRISPR abzielen
Viren, die Bakterien infizieren — Bakteriophagen — haben „Anti‑CRISPR“‑Proteine entwickelt, die diese Abwehr auf fast jeder Stufe sabotieren, vom Blockieren der DNA‑Bindung bis zum Verkleben der Schnittstellen von Cas9. AcrIIA7 gehört zu einer Familie, die dafür bekannt ist, Cas9 anzugreifen, doch war unklar, wie genau es wirkt. Die Autoren untersuchten AcrIIA7 aus einem Darmbakterium, Phocaeicola dorei, bestimmten seine dreidimensionale Struktur und testeten sein Verhalten in Lösung. Sie entdeckten, dass vier AcrIIA7‑Moleküle zu einem kompakten Tetramer mit drei Domänen zusammenfinden, einschließlich einer flexiblen „Kopf“‑Region. Die Oberfläche dieses Tetramers weist eine tiefe, positiv geladene Rinne auf — eine einladende Tasche für negativ geladene Nukleinsäuren wie RNA.
Ein Protein, das RNA greift statt Cas9
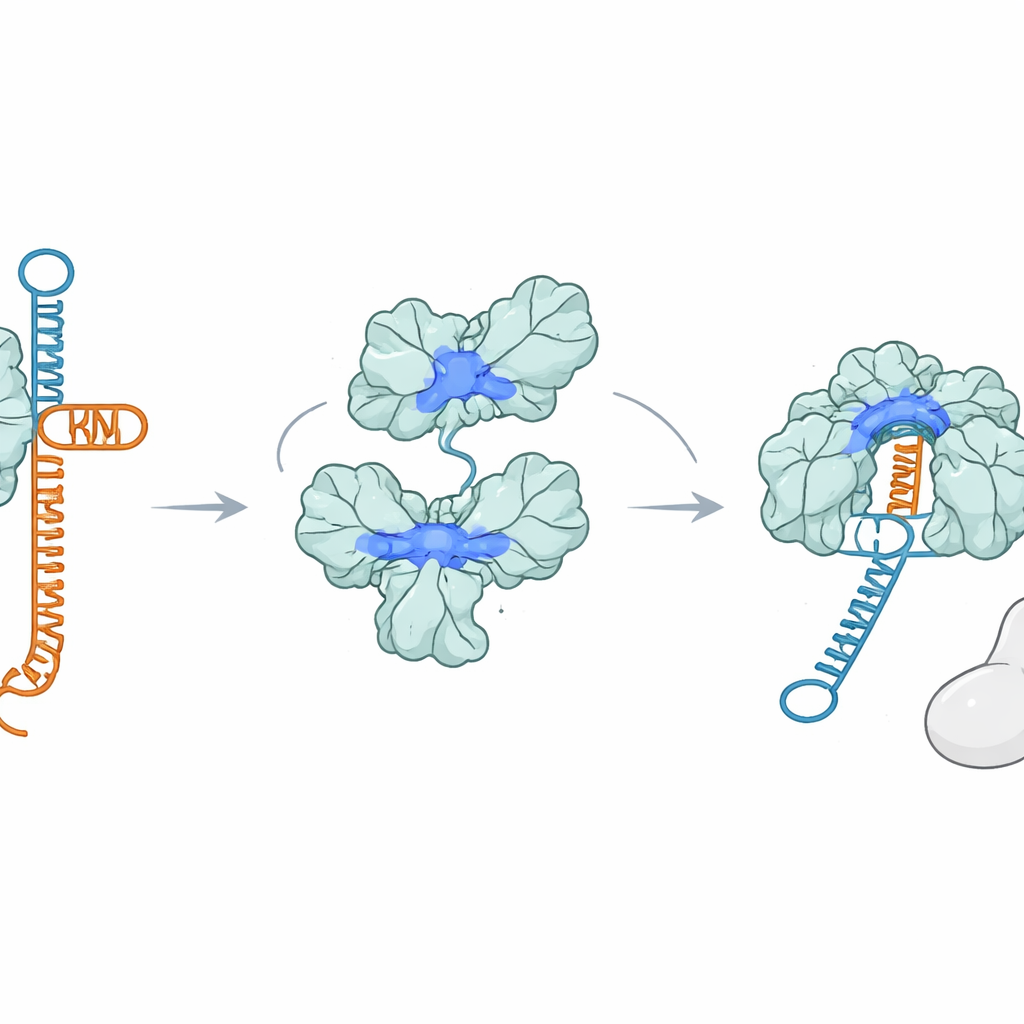
Überraschenderweise heftet sich AcrIIA7 überhaupt nicht an Cas9. Mithilfe mehrerer Bindungstests stellten die Forscher keine stabile Wechselwirkung zwischen AcrIIA7 und Cas9 fest, ebenso wenig mit einzelnen Teilen von Cas9. Stattdessen fanden sie, dass AcrIIA7 RNA direkt und selektiv bindet. Es ignorierte einzel‑ und doppelsträngige DNA, setzte sich aber stark an tracrRNA fest, eine der beiden RNAs, die normalerweise paaren, um die Anleitung zu bilden. AcrIIA7 erkannte nicht die exakten RNA‑Basen, sondern eine bestimmte gefaltete Form aus zwei benachbarten Haarnadel‑Schleifen. Wurden diese Schleifen entfernt oder die Gesamtarchitektur der Stängel‑Schleife verändert, schwächte oder verschwand die Bindung. Das zeigte, dass AcrIIA7 die Struktur der RNA mehr liest als ihre Sequenz.
Das Kapern des Leitfadens, bevor er entsteht
Indem es sich an tracrRNA klemmt, verhindert AcrIIA7, dass tracrRNA mit crRNA paaren kann, um den für Cas9 nötigen Leitfaden zu bilden. Versuche im Reagenzglas zeigten, dass, wenn AcrIIA7 der tracrRNA vor Cas9 begegnet, die nachfolgenden Schritte ins Stocken geraten: Die vollständige Leit‑RNA bildet sich schlecht und Cas9 kann die Ziel‑DNA nicht effizient schneiden. Sobald jedoch ein kompletter Cas9–RNA‑Komplex gebildet ist, kann AcrIIA7 diesen nicht mehr verdrängen, was unterstreicht, dass seine Hauptwirkung früh am Schritt der Leitfadenbildung liegt. Mutationen positiv geladener Aminosäuren, die die Rinne des Tetramers säumen, schwächten sowohl die RNA‑Bindung als auch die Cas9‑Hemmung, was die Vorstellung stützt, dass tracrRNA beim Kapern in diese Rinne einschnürt.
Zwei Varianten desselben Saboteurs
Das Team verglich außerdem das voll‑längige AcrIIA7, das die flexible Kopfdomain enthält, mit einer kürzeren, natürlich vorkommenden Variante ohne diesen Bereich. Beide Formen assemblierten sich zu Tetrameren und konnten tracrRNA sowie sogar kleine zyklische Signalmoleküle binden, die in anderen Immunwegen verwendet werden. Doch nur die kopflose Variante blockierte Cas9 stark, wenn bereits eine vollständig gebildete Leit‑RNA vorhanden war, was darauf hindeutet, dass die Kopf‑Region den Zugang zu längeren RNA‑Strukturen zum Teil behindert. Folglich stört das voll‑längige Protein hauptsächlich den frühesten Schritt — das Wegfangen freier tracrRNA —, während die kürzere Form den reifen Leitkomplex außerdem effektiver binden und blockieren kann.
Was das für Biologie und Genbearbeitung bedeutet
Für Nicht‑Spezialisten ist die Kernbotschaft, dass dieses virale Protein CRISPR nicht dadurch ausschaltet, dass es die bekannte Cas9‑„Schere“ direkt angreift, sondern indem es deren RNA‑Gerüst stiehlt, bevor die Schneidemaschine aufgebaut ist. Das offenbart einen neuen Schwachpunkt in CRISPR‑Abwehrmechanismen: die RNA‑RNA‑ und RNA‑Protein‑Kontakte, die zur Assemblierung eines funktionierenden Komplexes nötig sind. In der Natur hilft eine solche Strategie Viren, an bakterieller Immunität vorbeizuschlüpfen. Im Labor könnten von AcrIIA7 inspirierte Moleküle präzise, reversible Wege bieten, CRISPR‑Werkzeuge durch Anvisieren ihrer RNA‑Leitfäden abzuschalten und so die Sicherheit und Kontrolle künftiger Genom‑Editierungs‑Therapien zu verbessern.
Zitation: Lee, S.Y., Park, H.H. AcrIIA7 hijacks tracrRNA to block CRISPR-Cas system. Nat Commun 17, 3959 (2026). https://doi.org/10.1038/s41467-026-70749-w
Schlüsselwörter: CRISPR‑Cas9, Anti‑CRISPR‑Proteine, tracrRNA, Bakteriophagen, Kontrolle der Genbearbeitung